
 SoundEagle
SoundEagleThose who are interested in contributing photos or videos can upload them to the Queensland Orchid International Facebook Group.
Excellent or exceptional photos and videos uploaded to the group may be featured in the Gallery on Page 2 of this post to provide exemplary visual documentations of Flower-Pollinator Relationship and Insect-Plant Relationship.

Bumblebees and the flowers they pollinate have coevolved so that both have become dependent on each other for survival. (Photo credit: Wikipedia)
Coevolution can occur at many biological levels: it can be as microscopic as correlated mutations between amino acids in a protein or as macroscopic as covarying traits between different species in an environment. Each party in a coevolutionary relationship exerts selective pressures on the other, thereby affecting each other’s evolution.… Evolution in response to abiotic factors, such as climate change, is not biological coevolution (since climate is not alive and does not undergo biological evolution).
(Text credit: Wikipedia)

In consolidating his ideas about natural selection, and in considering orchids as being “universally acknowledged to rank amongst the most singular and most modified forms in the vegetable kingdom”, Charles Darwin (1809–1882) was already making concrete observations on, and evolutionary links between, insects and plants in the first edition of his book published in early 1862:
The object of the following work is to show that the contrivances by which Orchids are fertilised, are as varied and almost as perfect as any of the most beautiful adaptations in the animal kingdom; and, secondly, to show that these contrivances have for their main object the fertilisation of the flowers with pollen brought by insects from a distinct plant.
Indeed, one can endeavour to trace the history of evolutionary biology and come to the recognition that Darwin’s book on orchids generated the impetus for all succeeding investigations into coevolution and the evolution of extreme specialization. The book was published in 1862, less than 30 months after the advent of On the Origin of Species, and its second edition in 1877, when the contemporary Victorian vogue for growing exotic orchids was well established in England and Europe.
Riding on the vogue intentionally or unintentionally, the book On the Origin of Species presents Darwin’s first detailed demonstration of the potency of natural selection, and demonstrates how multifaceted ecological relationships can result in the coevolution of orchids and insects. In other words, it explains how the relationship between plants and animals can produce beautiful and complex forms, including exaggerated phenotypes with unusual traits or extreme morphologies. A famous instance is documented on page 162 of the second edition: Darwin recounted his receiving from the distinguished horticulturist Mr James Bateman (1811–1897) in 1862 a Madagascar orchid named Angraecum sesquipedale whose nectary was 11.5 inches long, and predicted that there must be a species of moth with proboscis long enough to reach the nectar at the end of the spur. It is no wonder that the orchid is given the specific epithet in Latin, “sesquipedale”, meaning “one and a half feet”. On page 165, Darwin even predicted that “[i]f such great moths were to become extinct in Madagascar, assuredly the Angræcum would become extinct.” Such a moth, named Xanthopan morganii, was discovered in 1903, validating the predictive power of (co)evolution.
 Commonly called Darwin’s moth, Darwin’s hawkmoth or Morgan’s sphinx moth, Xanthopan morganii is the species of the monotypic genus Xanthopan, which is classified under Sphingidae, a family of about 1,450 species of moths (Lepidoptera) commonly known as hawkmoths, sphinx moths, hummingbird moths and hornworms. Since nocturnal sphingids tend to be attracted to pale flowers with long corolla tubes and a sweet odour, a specific pollination syndrome has been named after the family of moths as “sphingophily”. Typical of sphingids, Xanthopan morganii has the ability to hover like hummingbirds and feed on the nectar of Angraecum sesquipedale’s flowers, which the sphinx moth identifies by scent. Then the moth flies backwards over 30cm and unrolls its mouthpart by pumping its haemolymph (a fluid plasma) into its coiled haustellum (a sucking organ or proboscis), which then uncoils and straightens under hydrostatic pressure before being inserted into the orchid’s flower, against which the moth has to press its head so that the tip of its haustellum can reach the nectar at the bottom of the flower’s long spur, causing the flower’s pollinarium to be stuck to the base of the moth’s haustellum.
Commonly called Darwin’s moth, Darwin’s hawkmoth or Morgan’s sphinx moth, Xanthopan morganii is the species of the monotypic genus Xanthopan, which is classified under Sphingidae, a family of about 1,450 species of moths (Lepidoptera) commonly known as hawkmoths, sphinx moths, hummingbird moths and hornworms. Since nocturnal sphingids tend to be attracted to pale flowers with long corolla tubes and a sweet odour, a specific pollination syndrome has been named after the family of moths as “sphingophily”. Typical of sphingids, Xanthopan morganii has the ability to hover like hummingbirds and feed on the nectar of Angraecum sesquipedale’s flowers, which the sphinx moth identifies by scent. Then the moth flies backwards over 30cm and unrolls its mouthpart by pumping its haemolymph (a fluid plasma) into its coiled haustellum (a sucking organ or proboscis), which then uncoils and straightens under hydrostatic pressure before being inserted into the orchid’s flower, against which the moth has to press its head so that the tip of its haustellum can reach the nectar at the bottom of the flower’s long spur, causing the flower’s pollinarium to be stuck to the base of the moth’s haustellum.

Figure 10. Xanthopan morganii praedicta visiting flowers of Angraecum sesquipedale (A–D) and Angraecum compactum (E, F). A, male moth positioned on the labellum during a visit. B, visiting moth flying up as it withdraws its proboscis which has a pollinarium (p) attached to near the base of its proboscis. The pollinia lie flat against the proboscis because their stipes are parallel to it. C, female moth with laterally positioned pollinia on the tip of a labellum. D, a male moth flying in an enclosure immediately after its capture. The tongue bears a viscidium (v) near its base and a remnant (n) of what is probably nectar 7.9 cm from the base. E, maximal insertion of proboscis just before the start of withdrawal. F, pollinaria (p) are removed during withdrawal (A–F, Wasserthal, 1997). Pollinia of Angraecum sesquipedale are transferred. Those of Angraecum compactum are not (Wasserthal, 1997).
Arditti, J., Elliott, J., Kitching I.J. and Wasserthal, L.T. (2012): ‘Good Heavens what insect can suck it’ – Charles Darwin, Angraecum sesquipedale and Xanthopan morganii praedicta. Botanical Journal of the Linnean Society, 169, 403–432. – Scientific Figure on ResearchGate. Available from: https://www.researchgate.net/figure/Xanthopan-morganii-praedicta-visiting-flowers-of-Angraecum-sesquipedale-A-D-and-A_fig10_262934621 [accessed 13 Feb 2018]

Closely related to Angraecum sesquipedale is Aerangis ellisii, also known as Aerangis platyphylla, which is not only ephiphytic but also epilithic, as it is often found to be growing on the surface of rock. It is a smaller orchid endemic to central and eastern Madagascar at elevations of 1100 to 1400 meters. As shown in the photo below and according to the abstract of a 1988 paper entitled “Hawk-moth scale analysis and pollination specialization in the epilithic Malagasy endemic Aerangis ellisii (Reichenb. fil.) Schltr. (Orchidaceae)” authored by L A Nilsson and E Rabakonandrianina:
The flowers exhibit long nectariferous spurs indicative of hawk-moth pollination, the mechanism evidently involving pollinia transfer via the frons and palpi of the vectors. Analysis of hawk-moth scales on naturally pollinated stigmas showed that the principal pollinators were Agrius conuolvuli (Linnaeus) and Panogena lingens (Butler). Proboscis morphology of long-tongued Malagasy Sphingidae in relation to nectar position and spur morphology in A. ellisii also indicated that A. conuolvuli and P. lingens were best suited to interact with the plant’s floral adaptation. Aerangis ellisii seems to display a relatively moderate specialization versus the Malagasy hawk-moth guild since its nectar is accessible even to those long-proboscis hawk-moth species which are not able to act as pollinators.

Four years later, in a 1992 paper entitled “Exact tracking of pollen transfer and mating in plants”, the abovementioned two researchers in conjunction with B Pettersson concluded on the coevolutionary outcomes of animal-plant interactions in pollination biology and plant morphology as follows:
UNLIKE animals, where individuals engage in direct sexual encounters, higher plants interact sexually only through minute, usually animal-mediated pollen grains, a trait that has hampered understanding of processes that govern plant evolution. A new technique using micro-tags to mark individual orchid pollinia and monitoring of all stigmas for pollination made it possible to measure exactly pollen transfer and mating pattern in a plant species. We report here that in populations of a hawkmoth-pollinated orchid, Aerangis ellisii, pollen transfers were found to be infrequent, to involve single pollen parents, and to occur mostly within 5 metres. Pollinator-mediated patterns of disproportional reproductive success suggest that floral traits are being shaped by mutual sexual selection as proposed by Darwin.
Even Angraecum sesquipedale’s smallest South African cousin, a miniature orchid called Mystacidium capense, is also exclusively hawkmoth-pollinated, its flower having a 3.9cm spur, much shoter than the 27–43cm (10.6–16.9in) spur of Angraecum sesquipedale. That coevolution so similar in kind has left its mark in species so dissimilar in size is quite remarkable. Taxonomically, both species had crossed paths at the level of genus, given that Angraecum sesquipedale was classified as Mystacidium sesquipedale (Rolfe) in 1904, whereas Mystacidium capense was originally called Angraecum capense (Lindl.) in 1830–40. Robyn P Luyt’s Master of Science Thesis of 2002 entitled “Pollination and evolution of the genus Mystacidium (Orchidaceae)” reveals that both Mystacidium capense and Mystacidium venosum are exclusively pollinated by long-tongued hawkmoths since their spur lengths are 3.9cm and 4.7cm respectively, whereas other Mystacidium species are pollinated by settling noctuid moths (and perhaps also bees, flies or even butterflies), since their spur lengths are less than 3cm, ranging from 1.68cm in Mystacidium tanganyikense to 2.56cm in Mystacidium gracile.

In the introduction to chapter 3 of the thesis on pages 33 and 34, Luyt also mentions Darwin and explains in the context of pollination biology that long spurs are evolved traits that confer “effective pollination by long-tongued pollinators” as the result of an evolutionary race between a plant and its pollinator:
The mechanism by which long spurs evolved was suggested by Darwin (1862) and later experimentally tested by Nilsson(1988): selection for longer spurs will compel a long-tongued pollinator to insert its proboscis deep into the spur, thereby ensuring contact with the flower’s sexual organs. Conversely, longer tongues are selected when they enable pollinators to reach deeply concealed nectar. The “evolutionary race” continues as pollinators with longer mouthparts drive the selection for longer spurs, and vice versa.
In biology, coevolution happens “when changes in at least two species’ genetic compositions reciprocally affect each other’s evolution.” Coevolution is considered as one of the most defining ecological and (phylo)genetic processes that organize biodiversity in ecosystems through interspecific interactions resulting from the relationship between individuals of two or more species in a community or biocenosis. According to John N Thompson, “coevolution is a pervasive process that continually reshapes interspecific interactions across broad geographic areas. And that has important implications for our understanding of the role of coevolution in fields ranging from epidemiology to conservation biology.” Regarded as a potent evolutionary force and feedback mechanism, the process of coevolution results in a series of adaptive changes in the genotypes and associated traits of the interacting species through inherent reciprocality of coevolutionary selection such that “the fitnesses of two interacting species depend not only on their own genotypes (and associated traits), but also on each other’s genotypes (and traits).”
Coevolution can also be called reciprocal evolution, a term encapsulating the process of mutual evolutionary change that occurs in pairs or groups of species interacting with each other and applying selection pressure to one another. Furthermore, coevolution can be deemed as an example or a subset of mutualism, where “two organisms of different species exist in a relationship in which each individual benefits from the activity of the other.” Specifically, pollination mutualism is a type of service-resource relationship in which “nectar or pollen (food resources) are traded for pollen dispersal (a service)”. Both coevolution and mutualism are thought to be significantly contributing to biological diversity in many ways, including flower forms (vital for pollination mutualisms) and coevolution between groups of species.
Angraecum sesquipedale and Xanthopan morganii as well as Mystacidium capense, Aerangis ellisii and the respective hawkmoths pollinating them, are three good examples of obligatory symbiosis or obligate mutualism, which is a form of symbiosis in which a close and long-term biological interaction between two different species is so indispensable or intricate that both species become highly interdependent and cannot survive on their own. Bound by this type of symbiosis, the host plant and the specific pollinator involved are called obligate mutualists or obligate species.
In contrast, a host plant whose pollination biology is less specialized tends to have non-exclusive nectaries accessible by multiple insect (or animal) species capable of pollinating its flowers. Such a host plant and its varied pollinators represent facultative symbiosis or facultative mutualism, a type of relationship where the survival of the host plant and the pollinator involved is not dependent upon the biological interaction, such that one facultative mutualist or facultative species can survive without the other and vice versa.
Overall, mutualism is essential for the health of many ecosystems, which would deteriorate or collapse without insect- or animal-mediated pollination and seed dispersal. Mutualism is believed to be highly responsible for both reciprocal specialization and coevolutionary diversification, to the extent that there are over one hundred times more existing flowering plant species (angiosperms, including herbaceous plants, shrubs, grasses and most trees) than there are species of all other extant seed-bearing plants (gymnosperms, including the conifers, cycads and ginkgo). In other words, more than 99% of seed plants are flowering plants, totalling over a quarter million species in more than 300 taxonomic families. Likewise, roughly 75% of all extant animal species are insects, the diversity of which is estimated to be from 2.6 to 7.8 million species with a mean of 5.5 million. That there are readily observable strong correlations between the number of angiosperm species, floral features and insect families is beyond any reasonable doubt. Appearing 130 million years ago, angiosperms became ubiquitous and diverse by 100 million years ago. The angiosperm radiation had been so rapid and saltational that the rise of flowering plants during the Cretaceous period was designated by Charles Darwin as the “abominable mystery” in his letter addressed to his friend Sir Joseph Hooker in July of 1879. However, the precise roles and contributions of pollinator-plant coevolution to the radiation and diversity of both insects and angiosperms require further refining to iron out the “empirical anomalies and theoretical inconsistencies” in the biotic pollination hypothesis. Moreover, the angiosperm radiation remains somewhat of an “abominable mystery” until more fossil evidence is available to adequately shed light on the rise of the flowering plants, which has also been linked to climatic change (such as increases in carbon dioxide from major geological events), the evolution of herbaceous plants, and even the coevolution with dinosaurs (the last of which was suggested by Bob Bakker in 1986, but yet to be proven with scientific evidence). Hopefully, advances in molecular genetics can contribute solid clues to, and promote more complete understanding of, the (co)evolutionary history of flowering plants and their pollinators.
In any case, insect- or animal-mediated pollination is a reproductive strategy that is not only vital to the evolutionary success and diversity of flowering plants, but also crucial to agriculture and human survival, as 75% of crop species depend on biotic pollination to disperse their pollens. Such evolutionary success and diversity are prodigiously reflected in the size of the Orchidaceae as well as the complex pollination mechanisms and fascinating pollination syndromes characteristic of the large family. Nevertheless, certain researchers in biology and environmental science propose that the taxonomic diversity of the Orchidaceae should not be overly or summarily “attributed to adaptive radiation for specific pollinators driven by selection for outcrossing”, since genetic drift, being one of the fundamental mechanisms of evolution (along with natural selection, mutation and migration) and driven by random environmental events or the vagaries of chance, invariably results in some species or individuals leaving behind a few more descendants (and thus their genes) than other species or individuals regardless of their fitness, to the extent that
orchids are primarily pollination limited, the severity of which is affected by resource constraints. Fruit set is higher in temperate than in tropical species, and in species which offer pollinator rewards than those that do not. Reproductive success is skewed towards few individuals in a population and effective population sizes are often small. Population structure, reproductive success and gene flow among populations suggest that in many situations genetic drift may be as important as selection in fostering genetic and morphological variation in this family. Although there is some evidence for a gradualist model of evolutionary change, we believe that the great diversity in this family is largely a consequence of sequential and rapid interplay between drift and natural selection.
The same researchers conclude that “pollen-limitation of both male and female reproductive success, in combination with the unique structure of the orchid flower (the column), largely explains the extravagant pollination mechanisms.… Paradoxically, it is the low reproductive success of orchids, not the opposite, that may account for both their unique pollination mechanisms and extreme diversity. The Orchidaceae is a remarkable group of plants and the fundamental processes by which the family has diversified are far more fascinating than Darwin ever suspected.” It is conceivable that the meagre reproductive success of orchids may have also contributed to their floral longevity, especially when successful pollination in the Orchidaceae is positively correlated to the flower lifespan, which is negatively correlated to pollinator visitation frequency, whether or not extreme specialization has been a contributing factor. In other words, many orchid flowers are long-lived because they have evolved to remain open and viable long enough to be visited by their pollinators, albeit on an infrequent basis. For a deeper understanding of the apparent discrepancies between floral forms and pollinator types, consult Dr Jeff Ollerton’s paper entitled “Reconciling Ecological Processes with Phylogenetic Patterns: The Apparent Paradox of Plant-Pollinator Systems”, written and published back in 1996, near the start of “[t]he explosion in orchids as houseplants”.
Whilst Darwin’s evolutionary prediction of the moth’s proboscis in 1862 is remarkable, this example of specific coevolution or pairwise coevolution (such as that of host and pollinator, host and symbiont, host and parasite, or predator and prey) is the exception rather than the rule, given that strict specialization of plants relying on one species of pollinator is relatively rare, as it can lead to variable or unreliable reproductive success when pollinator populations vary significantly or unpredictably. As with many forms of specialization, the exclusivity of a service-resource relationship via pollination mutualism can be a double-edged sword. Thus, plants usually generalize on a wide range of pollinators, and many insect pollinators are generalists visiting various types of flowers, insofar as such ecological generalization is ubiquitous in nature. However, some research “results suggest that a fine-tuned one-to-one coevolutionary state between a flower species and a pollinator species might be common, but frequently overlooked, in multiple flower-pollinator interactions”, and that in stable pollination systems, “one-to-one interactions are likely to be favorable, as opposed to the one-to-many or many-to-many interactions, typical in other mutualisms.” In contrast, some researchers not only caution against assuming that coevolution and selection pressures resulting from interspecific interactions necessarily or invariably lead to specialization, but also highlight the evolutionary advantages and wider ecological implications of plant generalization and pollinator generalization:
Pollinator generalization is predicted when floral rewards are similar across plant species, travel is costly, constraints of behavior and morphology are minor, and/or pollinator lifespan is long relative to flowering of individual plant species. Recognizing that pollination systems often are generalized has important implications. In ecological predictions of plant reproductive success and population dynamics it is useful to widen the focus beyond flower visitors within the “correct” pollination syndrome, and to recognize temporal and spatial fluidity of interactions. Behavioral studies of pollinator foraging choices and information-processing abilities will benefit from understanding the selective advantages of generalization. In studies of floral adaptation, microevolution, and plant speciation one should recognize that selection and gene flow vary in time and space and that the contribution of pollinators to reproductive isolation of plant species may be overstated. In conservation biology, generalized pollination systems imply resilience to linked extinctions, but also the possibility for introduced generalists to displace natives with a net loss of diversity.
The understanding of coevolution depends on the multifaceted investigation of community ecology (often simply called community, but also known as synecology or ecological community), which, according to Wikipedia, “is the study of the interactions between species in communities on many spatial and temporal scales, including the distribution, structure, abundance, demography, and interactions between coexisting populations. The primary focus of community ecology is on the interactions between populations as determined by specific genotypic and phenotypic characteristics. Community ecology has its origin in European plant sociology. Modern community ecology examines patterns such as variation in species richness, equitability, productivity and food web structure (see community structure); it also examines processes such as predator–prey population dynamics, succession, and community assembly.” Out of the study of community ecology came the theory of mosaic coevolution described as follows:
Mosaic coevolution is a theory in which geographic location and community ecology shape differing coevolution between strongly interacting species in multiple populations. These populations may be separated by space and/or time. Depending on the ecological conditions, the interspecific interactions may be mutualistic, antagonistic or even an arms race showing variation in specific traits over a broad geographical area. In mutualisms, both partners benefit from the interaction. When both partners experience a decreased fitness, it is an antagonistic relationship. Arms races consist of two species adapting ways to “one up” the other. Several factors affect these relationships, including hot spots, cold spots, and trait mixing. Reciprocal selection occurs when a change in one partner puts pressure on the other partner to change in response. Hot spots are areas of strong reciprocal selection, while cold spots are areas with no reciprocal selection or where only one partner is present. The three constituents of geographic structure that contribute to this particular type of coevolution are: natural selection in the form of a geographic mosaic, hot spots often surrounded by cold spots, and trait remixing by means of genetic drift and gene flow. Mosaic, along with general coevolution, most commonly occurs at the population level and is driven by both the biotic and the abiotic environment. These environmental factors can constrain coevolution and affect how far it can escalate.
The geographical mosaic theory was first described by Ehrlich and Raven in 1964 after studying butterflies that coevolve with plants. However, the idea of coevolution itself goes all the way back to Darwin.
Moreover, across the vast, open systems in the ecosphere, plants and animals tend to coevolve within extended networks of multispecies ecological interactions, resulting in diffuse coevolution or guild coevolution, in which whole groups of species interact with other groups of species, leading to changes that cannot be designated as instances of specific or pairwise coevolution between two species. Throughout the expanse of coevolutionary dynamics, new modes of flower-pollinator interaction and diverse forms of insect-plant relationship can emerge and evolve, even though some of them may initially elude discovery, confound expectation, or defy conventional classification. A good example can be studied in an academic article entitled “The Pollination Ecology of an Assemblage of Grassland Asclepiads in South Africa”, available from Annals of Botany, an international plant science journal publishing novel and rigorous research. It is also cited and contextualized by Wikipedia as follows:
… new types of plant-pollinator interaction, involving “unusual” pollinating animals are regularly being discovered, such as specialized pollination by spider hunting wasps (Pompilidae) and fruit chafers (Cetoniidae) in the eastern grasslands of South Africa. These plants do not fit into the classical [pollination] syndromes, though they may show evidence of convergent evolution in their own right.
All in all, phenotype variations emerge out of the coevolutionary processes within and between ecological networks, and phenotypes (as well as extended phenotypes) appear and disappear over time. The multispecies interaction patterns and coevolutionary dynamics are determined not only by biodiversity, the abundance of individuals expressing each phenotype, but also by sustainability, the preservation of viable environments in which individuals and species can persist with ongoing evolutionary change, and in which species interactions can be maintained. In this regard, there has been a substantial shift from preserving specific phenotypic variants by deploying taxonomic data to describe the constituent species through the traditional assessment of their morphological characters, to preserving the ecological and evolutionary processes via which biodiversity is engendered. In essence, understanding these processes with respect to coevolution is essential for improving the aims, scopes and outcomes of conservation. And in turn, the concept of species as having the objective reality or embodiment of gene flow, and the assumption or assertion that species is always the preferred unit and fundamental currency of conservation, can be reassessed. After all, conserving species through species-oriented conservation programmes is often futile and fraught with recurrent crises when the species being conserved are increasingly deprived of viable habitats or ecosystems, and thus cannot be sustainably settled and protected; whereas conserving habitats or ecosystems is more desirable, productive and holistic since it facilitates not only the protection of numerous species together but also the preservation of the processes and interactions amongst the species and the habitats or ecosystems.
There has also been a corresponding shift of the intellectual kind happening in the life of a featured author at ✿❀ Queensland Orchid International ❀✿ and a featured guest at 🦅 SoundEagle : Dr Craig Eisemann is a retired biologist and entomologist who used to work for a major government organization to conduct entomological research with respect to parasite-host relationship as applicable to molecular biosciences, molecular animal genetics, and agricultural sciences in general, and more specifically to tropical animal production, tropical agriculture, insect biochemistry and immunological control, such as vaccination and antibody-mediated inhibition. Like most researchers, Dr Eisemann’s career was occupied with projects with sharply defined problems, which he and his colleagues investigated and solved with the experimental method (though researchers in other related or unrelated fields may otherwise elect to use the historical, descriptive, correlational and/or causal-comparative methods, depending on academic disciplines, research problems and relevant contexts), in addition to dealing with administration, bureaucracy and corporate culture as they arose. Unlike Charles Darwin who lived in the age of the “gentleman scholar” and enjoyed substantial financial inheritance and professional freedom to pursue wide-ranging interests from biology to geology, modern-day academics and researchers are increasingly specialized or vocationalized, blindingly honing their skillsets on pinpointing minutiae to outshine others in their respective microniches. Gone are the big narratives and grand syntheses, unless one has the time, fortitude and resources to become a maverick pursuing truly revolutionary research or going against prevailing trends to wield long and meandering strokes on the large canvass of a book (such as Darwin’s 365-page The Various Contrivances by Which Orchids are Fertilised by Insects), let alone a watershed multi-chapter magnum opus (as exemplified by Darwin’s 502-page On the Origin of Species). In this respect, James Stapley, a marine biologist and orchid enthusiast working in the ASCLME Project for the United Nations Development Programme, has summed up the predicament and impasse whilst guest-blogging for Linda Markovina, a freelance photography and travel journalist, about “Darwin and the Lost Art of the Naturalist”:
From an age when a naturalist would study anything and everything of interest, we now have researchers who might spend their entire career working on so small a topic as a single species or even a single enzyme system. Whilst such a depth of knowledge can be absolutely invaluable, many of the “important” questions, particularly at a societal level, are really “big picture” or “synthesis” overviews of not only many different scientific disciplines, but also benefit from the input of social, economic, political and even “humanities” disciplines. In an era where we have so many specialists, most of whom can scarcely understand the jargon of the research group down the corridor, where are the generalists who can help us [to] pull together these separate threads and weave them into a richly informative tapestry?
Even the publication style that the modern research “industry” seems to encourage increasingly drives this fragmented approach. There is little room for the grand treatise or synthesis (the odd exception, like IPCC reports, aside) – modern publication is overwhelmingly dominated by many, very short, highly focussed papers, often many from the same research team on very slightly different aspects of effectively the same research question (and often from the same data). Publish or Perish, Research Output Volume (measured by number of publications – the accountant’s metric of scientific “productivity”) encourages more and more of this “mini-paper”, “same data, slightly different angle” approach.
This fragments the research landscape still further, with each paper more of a single pixel, rather than a complete photograph. Piecing together a coherent picture from all of those tiny dots of information might be likened to doing a gigantic jigsaw puzzle with the back of the pieces, in the dark, where many of the pieces are duplicated (or missing), and where there are no straight edges to help you out!
Funding agencies generally like to see a depth of research (and occasionally strong citation of that research) on a particular topic, or obviously related research avenues. I strongly suspect many of them would not positively review funding requests from a researcher with as varied a research output as a modern day Darwin. This perhaps suggests that “generalist” scientists will never be those that attract high “career researcher” ratings. Perhaps they’ll be more likely to lurk on the scientific “fringe” – people like science bloggers, journalists, and “hobby” researchers. Some of them might ultimately collate a lifetime of observations into one or more significant books – but this seems a remote possibility.
Furthermore, the pressure of having to satisfy research quotas or agendas, and the stress to fulfil corporate expectations and business interests, can create or exacerbate issues about integrity, transparency and reproducibility of scientific research, and also increase the likelihood of questionable research practices involving fraud, misconduct, misrepresentation and falsification, often in an effort to obtain some desired outcomes. One can no longer automatically assume that those who have been conscientious and ethical in applying research can remain untainted or unaffected by such practices, since retractions of scientific papers and the reasons for their retractions are not always publicized. Given that the transparency of the retraction process can be dubious, other researchers as well as the media and the public unaware of the retractions may quote or broadcast invalid research findings, or even make decisions or commitments based on invalid results.
In spite of this, those who are conscientious would still like to be confident that their due diligence can be exercised to foster good understanding about various research methodologies and pitfalls, including the art and science of falsification (and of questioning), in order to gauge the validity and reliability of research findings, including their interpretations and assumptions. These abilities are not necessarily easy to cultivate by being (or functioning as) a “normal” or “regular” academic, given that the science and philosophy of research (and of knowledge) are very complex, and there are good reasons to be so. Perhaps such abilities are even more essential in analyses of, or discussions on, subject matters that are seldom or inadequately explained, debated or resolved.
The contemporary tertiary education system and some of the intramural politics and bottom-line approaches operating at universities can be detrimental and even hostile to academic research, especially multidisciplinary undertakings. There are both limits and segregations imposed on intellectual liberty with respect to research territories and knowledge demarcations, beyond which there are barely scant platforms and rare opportunities to generate some debates, discussions or studies on issues never, seldom or inadequately debated or contested before. Since demonstrating a substantial engagement with existing literature and critiquing contested areas of thought are enshrined and mandatory, those areas that have not been researched or contested would tend to be much less favoured or noticed, if not automatically consigned to or considered as non-academic or second-rate materials. In other words, there are significant barriers to becoming maverick and holistic in one’s academic life, and to functioning as an exemplary liberal scholar, in addition to the added risk of being ignored, abandoned, consigned or ostracised as anachronistic proponents or promulgators of erudition and education. It is unfortunate that those who conduct research at colleges, universities or other tertiary institutions face constant pressure to have clearly defined research topics and agendas, which are supposed or expected to be concentrated on and shaped by the most contested, recognized or commensurable areas that comply or align well with the prevailing paradigm, academic climate and intellectual zeitgeist, in which ideas, data, models, methodologies and theories are created, examined, refined and fought over and over again by peers and rivals, and repeatedly quoted with zest by fellow scholars and aspiring students to show that they are up there with the most consequential leaders. Contests are usually fought contiguously, with rivals coming from within a discipline and focussing on specialism and micro-topics. At the multidisciplinary level, the dins and roars of such contests are few and far between, as fault lines seldom straddle continents of knowledge. Even when they do, they are often dismissed, misunderstood or ignored rather than examined or contested. The upshot is that academic research is increasingly reduced to playing an intramural game for “points” that earn coterie repute and disputable expectation, where conditions conducive to unbridled, exploratory or revolutionary ways of conducting research or investigation have become very illusive, even more so as colleges and universities opt to operate under the model of mass education and customer satisfaction, all too often underfunded, overburdened, vocationalized and instrumentalized, becoming more performance-managed, metricized, casualized and marketized under the pervasive influence of privatisation, consumerism, audit culture, managerialism and neoliberal orthodoxy.[❆] In a blog post entitled “The Imperative to Leave Academia”, Bharath Vallabha highlights the pressing issues and mounting challenges created by the ongoing incompatibility between the ancient form of elite academia and the modern aim of mass education and professionalization, as well as the urgent need for establishing grassroots changes outside academia.
The ancient form of academia is now buckling under the weight of the aims of mass education. For two hundred years, from the middle of the 18th to the middle of the 20th centuries, the ancient form and the modern aim coexisted relatively harmoniously. But in the last fifty years, academia’s egalitarian aim has been pushing against and breaking its ancient form.
… the threat is much deeper: most professors teaching the classes no longer have the privilege to be liberal scholars. This is not due to nefarious reasons of scientism or neo-liberalism or identity politics. Those are symptoms exacerbated by the simple – and now irreversible – fact of mass college education, and the pressure that puts on how jobs are allocated.
The situation will get worse over time. In order to bridge deep disagreements, the kind of holistic vision that liberal scholars provide is needed. But professionalization pulls exactly in the opposite direction. Instead of reconciling disagreements, it reinforces them by legitimizing boundary policing.…
… Professionalization has made it hard for professors to debate each other just as thinkers, as liberal scholars. But professionalization on its own, without holistic thinking, can’t determine how competing forms of expertise should be reconciled. The result is instead of debating ideas, a lot of effort goes into positioning oneself as the right kind of expert and the other as the wrong kind of expert.
This means that as society struggles with questions of expertise vs democracy, academia is not able to stand above the fray. Conservatives who want to shut down humanities departments or don’t believe in climate change support their views by saying, “Some experts agree with us!” the implication being that reason itself is partisan. Academics claim they are not partisan and can lead the way. But it’s unclear how they can lead the way when professionalization has splintered even philosophy into dozens of competing sub-specialties.
What is the alternative for academics? Give up professionalization? Not possible, since there is no other method for divvying up jobs. To have at least some academics be liberal scholars and so aim beyond specialization? This is the inevitable future. As more and more teaching is done by adjunct professors, and as public universities and small colleges are under increased economic and social pressures, it will be mainly the faculty at rich, private universities who will have the luxury to be undergraduates for life.
Society needs liberal scholars. People who think big and write big, who look for large patterns across whole fields of knowledge and the human condition. Who might fall on their faces and get a lot wrong, but whose risks might also pay off big or just be thought provoking. As academia becomes more professionalized, people struggling or unhappy in academia have a choice. They can leave to continue a life of liberal arts reflection outside academia, and so contribute in a grass roots way to creating a more reflective society.…
The coming digital age won’t accommodate the habits of the medieval or the industrial age university. It will disrupt them in unpredictable and far reaching ways, changing both academia and society in the process. If the disruption to academia ends up being very great and if we don’t prepare for that future by creating new possibilities outside, future generations will wonder how we could have been so complacent.
Taking all of the aforementioned matters into account, and in appreciating or even assuming the beneficial role and benevolent spirit of a liberal scholar in the digital age, one can begin to realize that this extensive post is much more than an attempt or an opportunity for a scientist (or anyone for that matter) to ask or broach the question “Do Plants and Insects Coevolve?”, a (kind of) question that cannot always be well served or properly approached by the format and scope of a conventional scientific article or experiment, especially when liberal scholars are considerably hamstrung by what and how they can research and publish in learned society organizations and educational or academic institutions. Indeed, whatever or however one would like to designate what can be amply witnessed and experienced here — a flashy webpage, a hybrid blog post, a topical assignment, an educational article, an academic paper, a literature review, or a dual-author essay — one could have qualitatively identified the pairwise coevolution, reciprocal evolution, mutualism, symbiosis and service-resource relationship between Dr Craig Eisemann and 🦅 SoundEagle, all of which are rendered even more plausible or desirable when one also considers the rationale and validity of the latter’s following statement about this website being a garden of art, science, poetry, music, video, graphics, cartoons, animations, games and puzzles in a beautiful, dynamically syndicated press containing posts, buzzes, events and social media updates commensurate with the aims and missions of 🦅 SoundEagle, whose tagline is “Where The Eagles Fly . . . . Art Science Poetry Music & Ideas”:
In turn, these fields and disciplines can become both catalysts and arenas for recognizing, (re)creating and (re)contextualizing various pursuits and domains with multiple reference points as a means of reflecting and communicating the current state of community and society, and as a way of fostering mutuality, reciprocity and complementarity.
Understood in these terms, the value of certain fields and disciplines can truly begin to be appreciated and defined beyond the straightforward results and immediate implications of admiring, studying, developing and improving them, to the extent that partaking in these fields and disciplines becomes not just an attempt at, or a process of, accruing and capturing knowledge, but also a fruitful endeavour to use certain pursuits and domains and their myriad embodiments and representations as a means of expressing ideas, which are themselves products of instinct (nature), experience (nurture) and technology (tools), guided and shaped by principles and forms of knowledge.
Whilst this highly protracted, tripartite and multilateral post is presented as a question in its title, it is by no means constituted as a kind of debate based on opinion, rhetoric or generalization. In contrast, the issues raised by the question are complex ones requiring scientific interrogation based on the gathering, analysis and review of multiple data as well as the assessment of interpretations of those data to do a topical post based on such a question sufficient justice in structured detail. For further readings and references, please consult the detailed list of Related Sites and Articles compiled by 🦅 SoundEagle at the end of this post.
In completing this Foreword, and before serving the ✿ Main Course ✿, 🦅 SoundEagle would like to invite all and sundry to contemplate the significance and profundity of Jason D Hoeksema’s opening statements about Geographic Mosaics of Coevolution:
Species constantly engage in strong interactions with other species — parasites, predators, prey, and mutualists. As a result, their traits may coevolve and diversify in geographic mosaics.… Since Darwin and Wallace, studies on coevolution have shown that species interactions can drive rapid and sustained evolutionary change in species at multiple spatial and temporal scales, generating genetic diversity within populations, leading to adaptive differentiation among populations, and often leading to ecological speciation (Schluter 2009). It has been argued that much of the diversity on earth is a consequence of coevolutionary diversification in species interactions (Thompson 1994, 2005, Ehrlich & Raven 1964). Clearly, studies of coevolution in species interactions can lend insight into the fundamental processes generating and maintaining biodiversity, including genetic and phenotypic diversity within and between species.
http://www.gettyimages.com/detail/114858663
Orchid (Dracula lafleurii) flower with pollinator flies, Ecuador
All of nature understands the rules,
the necessary common heartbeat,
the spark that lights the fuel
that rules the world.
The butterflies, the bees,
flowers and trees,
man and beast-
why can’t we
see?
Coexistence
Cooperation
Cohesion
Companionship
Co Creation-
Or it all falls away…….
Insects were among the earliest terrestrial herbivores and acted as major selection agents on plants. Plants evolved chemical defenses against this herbivory and the insects, in turn, evolved mechanisms to deal with plant toxins. Many insects make use of these toxins to protect themselves from their predators. Such insects often advertise their toxicity using warning colors. This successful evolutionary pattern has also been used by mimics. Over time, this has led to complex groups of coevolved species. Conversely, some interactions between plants and insects, like pollination, are beneficial to both organisms. Coevolution has led to the development of very specific mutualisms in such systems.
(Text credit: Wikipedia)

Many insects including hoverflies and the wasp beetle are Batesian mimics of stinging wasps. Two wasp species and four imperfect and palatable mimics. (A) Dolichovespula media; (B) Polistes spec.; (C) Eupeodes spec.; (D) Syrphus spec; (E) Helophilus pendulus; (F) Clytus arietes (all species European). Of note, species C–F have no clear resemblance to any wasp species. The three hoverfly species differ in the shape of their wings and body, length of antennae, flight behaviour, and striping pattern from European wasps. One fly species (E) even has longitudinal stripes, which wasps typically don’t. The harmless wasp beetle does not normally display wings, and its legs do not resemble those of any wasps. (Photo credit: Wikipedia)
ೋღஜஇ  CRAIG EISEMANN at SoundEagle🦅 இஜღೋ
CRAIG EISEMANN at SoundEagle🦅 இஜღೋ
Abstract
Coevolution is usually defined as a reciprocal influence of at least two interacting species on each other’s evolution. For insect–host plant interactions, possible examples include the development of toxic or at least distasteful chemical constituents in plants as an evolutionary response to insect attack and the evolutionary response of some plant-feeding insects to these, and various adaptations found in flowers and pollinating insects which combine to permit and enhance a mutually beneficial relationship. It remains a matter for debate whether such developments truly exemplify coevolution as defined above; the evidence in favour of this interpretation appears most persuasive for flowers and their pollinators and at least some symbiotic relationships between ants and plants, in view of the multiple apparent adaptations seen on both sides of these relationships. In some instances, such as the development of long nectaries in flowers and correspondingly long sucking tubes in their specialized pollinators, attempts to model the evolutionary processes involved mathematically may strengthen the evidential basis for deciding whether a true coevolutionary process has occurred. A presumed coevolutionary relationship between two species or groups of species may be extended by the intrusion of other species not originally involved in the relationship, as in the case of aggressive flower mimicry by predatory insects that prey on pollinators of the mimicked flowers. Another kind of extension of a putative coevolutionary relationship is exemplified by Mullerian mimicry, in which insect species that have evolved to tolerate and sequester toxic or distasteful chemical compounds from their food plants have also coevolved to resemble each other visually, and yet another by Batesian mimicry, in which a palatable species, which does not feed on a toxic host plant, has evolved to resemble visually a toxic insect that does. The association between plants and plant-feeding insects is one of great antiquity, and it is suggested that coevolutionary relationships in their broadest sense should be conceived as occurring between whole evolutionary lineages over long periods of geological time.
Introduction
Plants and the insects and other animals that interact with them can obviously each influence the ability of the other to survive and reproduce. For example, plant-feeding insects grow and develop by destroying a plant’s stem, leaf or root tissue, or its seeds, or else suck its sap, while insects that collect nectar or pollen from flowers may perform a crucial role in spreading pollen between plants (or individual flowers) and hence facilitate reproduction by plants of the species used in this way. Therefore, a clear potential exists for plants and the insects and other organisms associated with them to influence each other’s evolution, by selecting for traits that may confer some protection for plants against insect attack, facilitate insects’ exploitation of plants or enhance a mutually beneficial relationship. Some biologists have developed this concept further by suggesting that plants and plant-utilizing insects (as well as pairs of other interacting species) often cause reciprocal evolutionary change in each other, a process usually termed “coevolution”[1].
Insects and Plant Chemistry
The term “coevolution” was popularized in a well-known paper[2] by P R Ehrlich and P Raven, published in 1964, in which butterflies and the host plants fed on by their caterpillars were discussed as an example of mutual evolutionary change in two interacting groups of organisms. The authors demonstrated a broadly consistent pattern of association between taxonomic groups of butterflies and particular taxonomic groups of plants. Their discussion centred around the chemical constituents of the plants, in particular their “secondary” compounds. These are a complex mixture of various classes of chemicals, including alkaloids, terpenoids and glucosinolates, which are often produced in specialized epidermal cells and are commonly assumed to have no direct role in plant structure or physiology. The occurrence of these compounds varies widely among taxonomic groups of plants; many plant groups are characterized by the presence of particular classes of secondary compounds. These authors, like other investigators before them, suggested that the presence of secondary compounds evolved as a defence against plant-feeding insects, as many are known to be toxic, in some degree, to various organisms.
The argument presented in the above paper depended upon the concepts that insects feeding on plants do indeed exert significant “selective pressure” on them. In essence, the damage they cause significantly reduces the plant’s ability to survive and reproduce, and that the presence of secondary compounds is an evolutionary response of the plant to this pressure, conferring a degree of immunity to insect attack and hence improving the plant’s ability to survive and leave viable offspring. An “immune” plant species could then perhaps more easily colonize new habitats or ecological niches, giving rise to additional species in the process and resulting in the eventual appearance of a group of species, constituting a genus, family or even higher classification, characterized by the presence of one or more classes of secondary compounds. This process is sometimes described as “escape and (evolutionary) radiation” or “escape-and-radiate coevolution”.
Some insect species may in turn evolve to tolerate the toxic constituents of certain plants, often by evolving the capacity for biochemical detoxification of the compounds involved; this development would allow them to exploit these plants as food. The insects may in fact go further, evolving the ability to use these originally protective secondary compounds as “token” attractants (in the case of volatile compounds) or stimulants of feeding or egg-laying to assist their utilization of plants to which they have become adapted. Insects that have adapted to a plant species or group of species having a similar chemical constitution could then, in the absence of competition from other plant-feeding species, readily give rise to new species adapted to related plants having a generally similar, though not necessarily identical, suite of secondary chemical compounds. In this way, taxonomically-related groups of butterflies (and other plant-feeding insects) could come to use as their food plants, particular taxonomically-related groups of plants containing generally similar secondary compounds. The processes described above may be repeated with the appearance of novel plant defences (not only chemical; see below) and subsequent insect adaptations to them, resulting in a parallel proliferation of new plant and insect taxonomic groups. A hypothetical illustration of this is presented in the following figure.
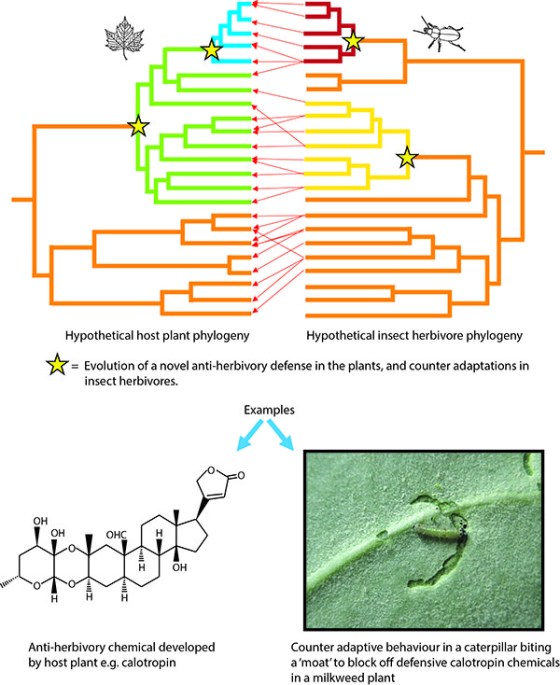
A hypothetical ‘escape and radiate’ example of coevolution, with a host-plant evolutionary tree, or phylogeny, on the left and an insect-herbivore phylogeny on the right. Red arrows: insect species feeding on a particular host. Stars: the origin of a novel anti-herbivory defence (in plants) or counteradaptations to overcome these defences (in insects). Different colours represent radiations with the new adaptations. Speciation occurs first in the plants and then in the insect herbivores (based on Ehrlich & Raven 1964 and Futuyma & Agrawal 2009).
Available from Fossil Focus: Arthropod–plant interactions (palaeontologyonline.com)
Slater, B. J. 2014. Fossil Focus: Arthropod–plant interactions. Palaeontology Online, Volume 4, Article 5, 1-17.
However, other researchers have argued that the above and other examples have not demonstrated conclusively the occurrence of coevolution, that is, of mutual evolutionary change in at least two interacting species, resulting from their interaction over time. For example, the Hungarian entomologist Tibor Jermy, in a series of papers published from 1976[3], contended that, in most natural ecosystems, plant-feeding (“phytophagous”) insects have a very much smaller biomass than their food plants and therefore inflict only minor damage on them. This means that the insects exert very little selective pressure on their host plants, so that there will be little tendency for the plants to evolve protective adaptations to their insect “predators”. Secondary chemicals, Jermy asserts, occur in plants because they perform functions unconnected with deterring herbivore attack — they may be intermediate metabolites or simply waste products of biochemical processes. Therefore, the process involved is not one of coevolution but of one-way “sequential evolution” — plants produce secondary chemicals for their own purposes and insects and other herbivores adapt to them.
A further objection to the idea that plant chemistry is, in part, an evolutionary response to insect attack is that many non-host plants are not in fact toxic to many phytophagous insect species[4] and even specific chemicals known to deter feeding and egg-laying may not be toxic[5], at least at the concentrations occurring in plants: insects’ host plant specificity is determined not by avoidance of toxicity but by other factors. These may include the presence or absence of sensory cues such as constituent volatile or non-volatile chemical compounds, and visual and tactile characteristics, all of which can mediate acceptance or avoidance of particular plant species. The original selective pressure that led insects to specialize in using certain host plants may have come from factors such as competition with other plant-feeding species and avoidance of predators[6], and the responses to specific sensory stimuli evolved by insects in identifying and locating their host plants may then have the effect of limiting them to host plants having the required presence or absence of sensory cues, irrespective of the potential suitability of other plants as sources of food[5].
These objections may be answered, to some degree, by noting firstly that even relatively slight selective pressure can, over many generations, result in significant evolutionary change[6] and secondly that patterns of utilization of plants by insects that are in evidence at present are the product of an evolutionary association between the two that has continued for tens or even hundreds of millions of years[7]. During this long association, many insects may have evolved to acquire a tolerance to many originally toxic plant chemicals that appeared as defences against insect attack in the remote past and persist, for various reasons, in a wide variety of present-day species. In any case, the fairly high degree of host-plant specificity observed in present-day insects will have the effect of limiting the abundance of most phytophagous insects because of the difficulty of finding suitable host plants scattered among many others in the ecosystem during the (usually short) lifetime of a searching insect. Such a constraint may have operated less strongly at an earlier stage in the evolutionary association between insects and plants when many insects may have been able to use a higher proportion of available plants, hence increasing the selective pressure on plants to evolve chemical and other defences. In any case, it is important to recognize that, as for other characters, plant chemistry may result from a variety of selective influences; even if defence against phytophagous insects and other herbivores is accepted as being important among these, it is not necessary to attribute the entire suite of plant secondary chemicals to this cause.
Other Plant Defences
Current plant defences are not limited to toxic secondary chemicals, and may include such characters as fine hairs (trichomes) on leaves and stems and latex circulating in plant conductive tissues that may immobilize and kill small larvae. An interesting example is the ocurrence of small yellow structures on leaves and stems of Passiflora species (passion vines) that resemble visually the eggs of butterflies of the genus Heliconius that feed on Passiflora as larvae. This apparent adaptation is illustrated in the following article.
http://www.passionflow.co.uk/downloads/gilbert_1982.pdf
These structures tend to deter the butterflies from laying eggs on these plants, as they normally avoid laying eggs near existing eggs, thereby minimizing overcrowding and cannibalism among larvae. This example may show evidence of a “coevolutionary arms race”, in which a plant has responded to insect attack by evolving toxic chemical constituents (in this case including cyanogenic glucosides) that deter most herbivores from feeding on them, a group of insect species has in turn evolved a biochemical tolerance for these compounds, allowing them to feed on an otherwise toxic plant, and the plant has in turn responded to this further challenge by evolving deterrent structures specific to these insects.
Insects and Flowers
A seemingly less controversial example of plant-insect coevolutionary relationships is provided by flowering plants and nectar- and pollen-feeding insects. Pollination of flowering plants by insects may have originated in early angiosperms, which were all initially wind-pollinated. It has been suggested that, as in extant gymnosperms such as conifers, droplets of sap were secreted by the ovule to catch pollen grains floating in the air. Insects began to use this sap as a food resource, incidentally transferring pollen grains between plants. As both “partners” benefitted from this relationship, natural selection acted on both groups of organisms to produce, on the one hand, more “attractive”, easily-located flowers, and on the other, specific structural, physiological and behavioural adaptations to allow the insects to better locate the flowers and to exploit their contained nectar and/or pollen. In general terms, this may constitute an example of “guild” or “diffuse” coevolution, which occurs between groups of species, rather than individual species. Different plant species may nevertheless evolve to target particular groups of pollinators or even individual species. For example, flowers pollinated by diurnal insects (those active during daylight) such as butterflies or wasps, which are often strongly visually-oriented, are frequently brightly-coloured, whereas flowers pollinated by nocturnal insects (such as most moths) tend to be pale-coloured and hence more visible in low light. Flowers visited predominantly by nocturnal foragers also produce more odourous chemicals, this characteristic being more suited to these insects, which tend to be more dependent upon responses to odours. Some additional information about flowers and pollinators is available on the following webpage.
http://biology.clc.uc.edu/courses/bio303/coevolution.htm

On the left, Angraecum sesquipedale, also known as Darwin’s orchid, Christmas orchid, Star of Bethlehem orchid, Madagascan Star orchid, Comet orchid, King of the Angraecums, and The One and a Half Foot Long Angraecum, is an epiphytic orchid endemic to Madagascar. (Photo by Michael Wolf).
On the right, Xanthopan morganii, also known as Morgan’s sphinx moth, is from East Africa (Rhodesia, Nyasaland) and Madagascar. (Photo by Esculapio).
A strikingly close evolutionary relationship between a single species (or group of closely-related species) of plant and a single species of pollinator is exemplified by the Madagascan Star Orchid Angraecum sesquipedale and its pollinator, the hawkmoth Xanthopan morganii.[19] In this orchid, the nectar is produced at the end of a remarkable corolla tube in the form of a spur some 20 to 35 cm long. In his book[8] on pollination of orchids, published in 1862, Charles Darwin predicted that some moth with a haustellum (specialized sucking tube) sufficiently long to reach the nectar at the end of this spur must exist in the orchid’s geographical range. Subsequently, Darwin’s contemporary and fellow evolutionary theorist Alfred Russel Wallace suggested that this moth would probably be a hawkmoth (family Sphingidae). In 1903, X. morganii, which was already known from Africa, was found to occur in Madagascar as well, but it was not until 1997 that it was shown to pollinate A. sesquipedale and other members of its genus. The orchid and its pollinator are illustrated below.
A video of the moth visiting this orchid in its natural environment can be viewed below.
In this mutualistic association between species, the coexistence of an unusually long nectary in the orchid and an unusually long haustellum in the moth has the effect of limiting pollination of these orchids to the one species of pollinator. This situation has the potential advantage for the orchid, which is said to be quite rare in its natural forest habitat, of ensuring that its pollen is transferred preferentially to other individuals of this species rather than being “wasted” by transfer to flowers of other species. Whilst having a long haustellum does not necessarily preclude access to nectar in species that do not have a long corolla tube, there may be an advantage to the moth (which is apparently also rare in its natural habitat) in specializing in collecting nectar from a particular species (or group of related species) for which it has no competitors, and hence a more assured supply of nectar. For the success of this relationship, it is important that the nectary be long enough to exclude other nectar-feeding species but not too long to be reached by X. morganii. Further, it is important for the orchid that the moth’s haustellum be only just sufficiently long to reach the nectar at the tip of the corolla tube, so that the head of the moth rubs against the anthers of the flower, collecting pollen that can subsequently be transferred to other flowers of the same species.
 Angraecum sesquipedale (Darwin’s orchid, Christmas orchid, Star of Bethlehem orchid, Madagascan Star orchid, Comet orchid, King of the Angraecums, The One and a Half Foot Long Angraecum), Madagascar
Angraecum sesquipedale (Darwin’s orchid, Christmas orchid, Star of Bethlehem orchid, Madagascan Star orchid, Comet orchid, King of the Angraecums, The One and a Half Foot Long Angraecum), Madagascar
Much commentary about the causes of particular evolutionary events is necessarily speculative, concerning as it does events and conditions that occurred in the remote past. In some cases, mathematical modelling may help to put such speculations on a somewhat firmer basis. Deep corolla tubes and long sucking tubes in pollinators appear to have evolved repeatedly in orchids and hawkmoths as well as in other groups of organisms, the pollinators including hummingbirds, nectar-feeding bats and certain species of flies. A group of researchers have attempted to model the evolution of these adaptations in general terms. Their models[9][10] have shown firstly that, in the presence of pollinators with long and short tongues that are competing for nectar, coexisting plant species will evolve corolla tubes of different lengths, as this will increase the proportion of their pollen grains being deposited on flowers of the same species. Secondly, for two plant and two pollinator species, all having a similar range of corolla tube depth or tongue length, variability in corolla tube depth within the two species will result in the evolution of different tongue lengths in the two pollinator species, provided that it is not equally costly in terms of energy for the two species to increase their tongue length, and that scarcity of nectar is a limiting factor for the survival of the pollinators. Once a difference in tongue length between the two pollinator species has become established, a divergence in corolla tube depth between the two plant species will develop. These conclusions hold for a wide range of assumptions about prevailing conditions, and therefore provide evidence that coevolution of these characters in interacting species can indeed occur. In explaining how “Resource Competition Triggers the Co-Evolution of Long Tongues and Deep Corolla Tubes”, Miguel A Rodríguez-Gironés and Ana L Llandres Resource demonstrate their main model components as follows.
Schematic representation of the main model components:
The foraging cycle (A) is iterated over 10,000 time units. Steps indicted in boxes with dark-blue outline require time, during which moths spend energy at a rate that increases with the length of their proboscis (as indicated in the box at the upper-left corner). The energy is recovered through nectar consumption (box with green background). The decision whether to exploit the flowers of a plant is probabilistic, and the probability of accepting a plant depends on the corolla depth of its flowers (box in the lower-left corner). When a moth exploits a flower, pollen can be transferred from the flower to the moth and from the moth to the flower, with different probabilities (B). At the end of the season, ovules are fertilised (C). The probability that a pollen grain fertilises an ovule depends on whether it arrived to the stigma early or late. Pollen grains from the same plant have a lower probability of fertilisation, and heterospecific pollen grains can prevent ovule fertilisation.
Ants and Plants
The associations that have developed between certain species of ant and particular plants provide further likely examples of coevolutionary relationships. More than 100 genera of plants are known to be “myrmecophytes”[11] — plants that live in a mutualistic relationship with a colony of ants. The plants provide food for their associated ants in the form of sugars and amino acids secreted from extrafloral nectaries or via sap-sucking insects that first obtain them from the plant, and in some cases in the form of specialized structures rich in proteins and lipids. Myrmecophytes also provide “domatia” — specialized plant structures containing cavities that can be occupied as nesting sites by their ant symbionts.
Several species of acacia (the “swollen thorn” acacias native to the tropical Americas) have evolved strong mutualistic relationships with ants of the genus Pseudomyrmex. The association between Vachellia (formerly Acacia) cornigera[12] and Pseudomyrmex ferruginea[13] has been studied in detail by Daniel Janzen[14]. This species of tree has evolved a number of adaptations, not shared with non-myrmecophytic acacias, that foster this relationship: enlarged thorns in which the ants find shelter (domatia), enlarged leaf nectaries from which the ants obtain sugars and amino acids, modified leaflet tips (“Beltian bodies”) rich in proteins and lipids that are consumed by the ants and their larvae, and year-round production and maintenance of leaves, even in dry seasons.
Occupation of a Vachellia cornigera plant by Pseudomyrmex ferruginea begins with a dispersing mated Queen attracted by the odour of the acacia. She then cuts her way into a hollow thorn or uses an existing entrance hole; inside the thorn she lays eggs and rears the first brood of workers. As the colony expands, the workers eventually occupy all of the thorns on the plant and sometimes spread to neighbouring plants of the same species; the population of a colony may reach as many as 30,000 workers. The ants, which are highly aggressive and possess, to human perception, an especially painful sting, patrol the host tree, killing most intruding insects, deterring browsing mammals and even pruning encroaching plants of other species to maintain a clear space around it. (However, the ants protect scale insects feeding on the host tree, as these are a source of honeydew consumed by the ants). The following illustration and video show some activities of P. ferruginea on its host tree:

The relationship is essentially an obligate one on both sides: the occupying ants obtain the great majority of their food from the tree and have adapted many aspects of their behaviour to occupy, exploit and protect their symbiotic plant; on the other hand, the benefits to the acacia from the association are shown by the fact that individuals of V. cornigera deprived of their ant symbionts are severely damaged by phytophagous insects and browsing goats and rodents, and in many environments tend to become overgrown by competing plant species. An interesting observation made by Janzen is that this species of acacia appears to have lost its ability to resist attack by insects and mammals as well as competition from other plants. Its leaves are not bitter to human taste, unlike those of non-myrmecophytic acacias, and are preferred to them by browsing mammals if the tree is not occupied by P. ferruginea. This species seems to have ceased using energy to produce feeding-deterrent chemical compounds; these are presumably not necessary since their protective functions have been assumed by the ant symbionts. Instead, V. cornigera has diverted energy towards producing outsized thorns, large extrafloral nectaries and Beltian bodies as supports for its myrmecine defence system.
A further example of a mutualistic relationship between a plant and an attendant ant species is provided by the carnivorous fanged pitcher plant Nepenthes bicalcarata[15] and a species of carpenter ant, Colobopsis (formerly Camponotus) schmitzi[16]. The pitcher plant inhabits mainly peat forests in north-western Borneo. N. bicalcarata is unique among some 120 species of its genus in being a myrmecophyte; it is also unusual in lacking a waxy layer on the peritreme (the “lip” of the insect-capturing “pitcher”), and in having a fluid inside its pitchers that is less acidic and viscoelastic than that of other Nepenthes species. In addition, it may lack the digestive enzymes that occur in the pitcher fluid of other species of its genus.
The associated ants nest in domatia formed in swollen, hollow tendrils; worker ants also conceal themselves underneath the peritreme, from where they “ambush” insects landing on the peritreme. They also appear to clean the peritreme of debris and fungal growth. The ants can swim actively in the pitcher fluid, from which they remove the larger prey items that fall into it, as well as some of the infauna of insect larvae (such as mosquito larvae) that normally inhabit the pitcher fluid. The low viscoelasticity of the fluid enhances the ability of the ants to remove prey from it, and the low acidity and possible lack or inactivity of digestive enzymes[17] permit both ants and infauna to survive in it. The next illustration and video show some of the details of the relationship.

The ants obviously consume part of the prey (and infauna) found in the pitchers; on the other hand, they benefit the plant by increasing the capture rate of prey through ambush and the preservation of a smooth peritreme surface, by digesting prey and making part of it available to the plant by depositing faeces into the pitchers, by providing ant carcasses which fall into the pitchers and by protecting the plant from phytophagous insects, in particular from a species of weevil that attacks growing shoots. They may also assist the plant by preventing accumulation of organic material in the pitchers to the point where putrefaction occurs, an event that would probably be deleterious to the plant, as well as to its possibly beneficial infauna.
In return, N. bicalcarata provides C. schmitzi with nesting sites in the form of domatia, prey occurring in and around the pitchers, sugar and amino acids from the large nectaries in the “fangs” that are located above the mouth of each pitcher. The net effect of the association on the plant seems to be definitely beneficial:[17] N. bicalcarata plants inhabited by C. schmitzi have more leaves, a greater total leaf area, less abortive pitcher development (as a result of protection from weevil attack) and larger pitchers. Plants colonized by C. schmitzi have been shown to have more nitrogen available to them than those not colonized, and more of this nitrogen is of insect origin.
The relationship for N. bicalcarata is, however, facultative rather than obligate, as the plant can survive in the absence of its ant symbiont. Nevertheless, plants without C. schmitzi colonies appear to derive no net benefit from their pitchers; the adaptations that foster the mutualistic relationship with this ant species (such as low acidity and viscoelasticity of its pitcher fluid and the possible lack of digestive enzymes) appear to have reduced its ability to derive nutrients from its pitchers in the absence of its ant symbiont. N. bicalcarata has adopted an alternative strategy for obtaining supplementary nutrients from its prey as compared with other species of its genus. For C. schmitzi, the relationship with N. bicalcarata appears to be an obligate one: it is totally dependent on its partner for nutrients and nest sites and apparently rarely leaves its plant host.
Extension of Coevolutionary Relationships
In some cases, an established interaction between two species or groups of species may be intruded upon by other species in their community which act as “free-riders” on a mutualistic relationship. An example is provided by the orchid mantis Hymenopus coronatus which has evolved to exploit the relationship between flowers and the various pollinators that visit them. These pollinator insects are preyed upon by the mantid when they are attracted to it because of its strong visual resemblance to flowers (in terms of insect perception, probably to a “generalized” flower rather than specifically to an orchid[18]), despite the strong visual similarity (to human perception) to some orchids that is shown on the webpage below.

A presumed coevolutionary relationship has therefore strongly influenced the evolution of an organism that was not originally part of the relationship. Whether the mantid has itself entered into a coevolutionary relationship with either the flowers or their pollinators (or both) is unclear, and would depend largely upon whether it exerts any significant selective pressure on them, that is, by preying on the pollinators and thereby having an incidental disruptive effect upon pollination.
A different kind of “third party” effect, in which a putative coevolutionary relationship has become extended to affect indirectly one or more additional species can be seen in mimicry involving toxic or distasteful insects. As discussed above, some insects have evolved to tolerate particular toxic secondary chemical constituents of plants, and hence can use plants containing these as food. Such toxic substances can in some cases be stored in the feeding insects and even transferred to later life stages. In other words, toxins accumulated by a caterpillar feeding on a host plant that is toxic to most other animals can be passed on to the later adult stage, such as a butterfly or moth that does not itself feed on the plant. Such insects may then be protected against predators by their stored toxic compounds.
Toxic or distasteful insects and other animals frequently display aposematic (warning) colouration, typically involving a prominent combination of black with yellow, orange or red. This is of advantage to the toxic animals as vertebrate predators, in particular, can learn to avoid prey showing this colouration after having a few unpleasant experiences with them. Often two or more such toxic species, which may or may not be closely related or feed on the same plant species, will evolve to resemble each other visually, so that fewer members of each species will be sacrificed in “educating” predators about the undesirability of eating them: a lesson learned from one such species will apply equally (or nearly so) to the other species in this “mimetic complex”. This condition is referred to as “Mullerian mimicry”. A striking example of this is provided by species of the tropical American butterfly genus Heliconius, the larvae of which feed on species of Passiflora containing highly toxic cyanogenic glucosides, and accumulate these compounds in their tissues and in those of subsequent life stages. The species H. erato and H. melpomene occupy similar geographic ranges in Central and South America, and both show remarkable variation in the colour patterns of their wings among subspecies occupying different areas within their ranges. As is shown in the figure below, there is a high degree of concordance in the wing patterns of subspecies of the two species that occur in the same areas: in each geograpical area, the locally occurring subspecies of these two unpalatable Heliconius species have evolved to resemble each other much more than either resembles other subspecies of its own species occurring elsewhere. In this and other cases, a presumed coevolutionary relationship between plants and phytophagous insects has led to a further coevolutionary relationship between the insects involved, with the effect of maximizing the survival benefit to them of storing plant-derived predator-deterring chemical compounds.

Heliconius erato and Heliconius melpomene resemble or mimic each other in many different countries, often sharing either bands or rays.
It may also occur that an insect that is not itself toxic or distasteful will evolve to resemble one that is, thereby acquiring some protection from predators (“Batesian mimicry”). In this case the mimic is a “free-rider” on the beneficial relationship that its model has developed with the latter’s host plant. Normally, the mimic is much less abundant than its model, so that the latter’s “lessons” to predators are not counteracted unduly by the presence of the palatable mimic.
 illustrating Batesian ...")
Plate from Bates (1862) illustrating Batesian mimicry between Dismorphia species (top row, third row) and various Ithomiini (Nymphalidae) (second row, bottom row) (Photo credit: Wikipedia)

Mimicry in butterflies is seen here on these classic “plates” showing four forms of H. numata, two forms of H. melpomene, and the two corresponding mimicking forms of H. erato. This highlights the diversity of patterns as well as the mimicry associations, which are found to be largely controlled by a shared genetic locus. (Photo credit: Wikipedia)
In some species of mimic, a similar effect is achieved through polymorphism, whereby different forms occur (often in the same geographical area) resembling different distasteful model species of widely differing appearance. A remarkable instance of this is provided by the African Mocker Swallowtail butterfly Papilio dardanus, the female of which has more than 12 different forms of wing shape and colour, several of which mimic diverse toxic species of butterfly. Some females of P. dardanus closely resemble the males of this species, which are not mimics. In the illustration in the webpage below, various model species from another butterfly family are shown above the line, and corresponding mimetic (and non-mimetic) forms of P. dardanus are shown below it.

Butterfly Genetics Group: Martin Thompson
Models are above the horizontal line, mimetic forms of P. dardanus are underneath.
Concluding Remarks on Coevolution
In answering the question “Do plants and insects coevolve?”, it is crucial to establish whether each partner in a relationship has influenced the evolution of the other[1]. In some instances, such as that of flowers and their insect pollinators, as well as that of certain plants and their ant symbionts, it appears highly probable that this has occurred. The presence of specializations such as, on the one side, visually conspicuous petals and the production of energy-rich nectar and specialized volatile chemical compounds, and on the other, of specialized mouthparts for obtaining nectar together with sensory and behavioural capabilities attuned to locating flowers using visual and chemical cues seems difficult to account for without invoking reciprocal evolutionary change. In other instances, such as that of plant chemistry, the picture is more equivocal: it is often difficult to prove that insect adaptations to the presence of particular chemical compounds are not purely unilateral ones to characters that evolved in plants for reasons unrelated to insect attack. Even the existence of a mutualistic relationship is not in itself sufficient evidence that a coevolutionary process has occurred; again, it is possible that characters in an organism that are of benefit to its partner may have evolved originally for reasons unrelated to this partner[1].
It is also important to recognize that many evolutionary adaptations may have occurred not in extant species but in ancestral forms (perhaps distantly ancestral) and have been preserved in their descendants through natural selection because of their continuing usefulness (perhaps not for the original reason). The biological agent that applied the selective pressure resulting in the original adaptation may not have been the same as or even directly ancestral to that which presently interacts with the species concerned[1]. Plants and phytophagous insects have coexisted and interacted over vast periods of time. The main insect orders (including grasshoppers, bugs, beetles, butterflies and moths, and wasps, bees and ants) that contain numerous plant-feeding species are known to have existed since at least the Jurassic period (circa 210 to 145 million years ago) and often considerably earlier than this[7], antedating the appearance of flowering plants. In considering plant-insect coevolution, it is necessary to take into account events occurring over deep time.
The causes of evolutionary events often remain speculative and controversial, having occurred mostly in the remote past, when conditions may have differed markedly from those prevailing at present. As a result, it is not surprising that it is often difficult to decide whether a true coevolutionary relationship has existed, especially for associations of great antiquity. For more recent associations, a strong indication of coevolution exists if both species or groups of species in an association show characters that are not shared by closely related species. The existence of exaggeratedly long corolla tubes in flowers and commensurately long sucking organs in their pollinators would appear to exemplify this situation. Other likely examples are provided by the physiological and behavioural adaptations on both sides of the mutualistic ant-plant relationships discussed above. Coevolution in its fullest sense, however, should perhaps be conceived as occurring over whole evolutionary lineages, including organisms not necessarily contemporary with each other, and not merely between pairs of extant species. The complexities of such considerations are likely to remain a matter for experiment and speculation by evolutionary theorists far into the future.
References
- ^ Janzen, D H (1980) When Is It Coevolution? Evolution 34: 611-612. Available at http://www.bio.miami.edu/horvitz/Plant-animal%20interactions%202013/coevolution/required%20readings/Janzen%201980.pdf
- ^ Ehrlich, P R and Raven, P (1964) Butterflies and Plants: A study in coevolution. Ecology 18: 586-608. Available at http://www.esf.edu/efb/parry/Insect%20Ecology%20Reading/Ehrlich_Raven_1964.pdf
- ^ Jermy, T (1976) Insect-Host Plant Relationship — Co-Evolution or Sequential Evolution? Symp. Biol. Hung. 16: 109-113
- ^ Common, I F B and Waterhouse, D F (1981) Butterflies of Australia (2nd Edition). Sydney and Melbourne: Angus and Robertson
- ^ Bernays, E and Graham, M (1988) On the Evolution of Host Specificity in Phytophagous Arthropods. Ecology 69: 886-892
- ^ Thompson, J N (1988) Coevolution and Alternative Hypotheses on Insect/Plant Interactions. Ecology 69: 893-895
- ^ Kulakova-Peck, J (1991) Fossil History and the Evolution of Hexapod Structures. in CSIRO, The Insects of Australia (2nd Edition). Melbourne: Melbourne University Press, pp. 141-179
- ^ Darwin, C (1862) On the Various Contrivances by Which British and Foreign Orchids are Fertilized by Insects, and on the Good Effects of Intercrossing. London: John Murray
- ^ Rodriguez-Girones, M A and Santamaria, L (2007). Resource Competition, Character Displacement and the Evolution of Deep Corolla Tubes. Am. Nat. 170: 455-464
- ^ Rodriguez-Girones, M A and Llandres, A L (2008) Resource Competition Triggers the Co-Evolution of Long Tongues and Deep Corolla Tubes. Available at http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0002992
- ^ Myrmecophytes https://en.wikipedia.org/wiki/Myrmecophyte
- ^ Vachellia cornigera https://en.wikipedia.org/wiki/Vachellia_cornigera
- ^ Pseudomyrmex ferruginea https://en.wikipedia.org/wiki/Pseudomyrmex_ferruginea
- ^ Janzen D.H. (1966) Coevolution of Mutualism Between Ants and Acacias in Central America. Evolution 20: 249-275. Available at http://www.jstor.org/stable/2406628
- ^ Nepenthes bicalcarata https://en.wikipedia.org/wiki/Nepenthes_bicalcarata
- ^ Colobopsis schmitzi https://en.wikipedia.org/wiki/Camponotus_schmitzi
- ^ Bazile V, Moran J A, Le Moguedec G, Marshall D J and Gaume L (2012) A Carnivorous Plant Fed by Its Ant Symbiont: A Unique Multi-Faceted Nutritional Mutualism PLoS ONE 7(5): e36179. Available at https://doi.org/10.1371/journal.pone.0036179
- ^ O’Hanlon J C, Holwell G I, and Herberstein M E (2014) Predatory Pollinator Deception: Does the Orchid Mantis Resemble a Model Species? Current Zoology 60: 90-103. Available at http://www.currentzoology.org/temp/%7B3E2BECB0-59F7-49BD-8B76-4D2B221A61E1%7D.pdf [Note: The link seems to be defunct.]
- ^ The spelling morganii was used in the original description of this species; however, the form morgani is also commonly encountered in the published literature. According to the International Code of Zoological Nomenclature, although it appears that the original spelling, involving as it does a non-Latin name, is not consistent with the current Code (Article 31), it should nevertheless be preserved (Articles 32 and 34). The spelling morgani is therefore incorrect (Article 33).
Keywords
Evolution, Coevolution, Insect-Plant Relationship, Flower-Pollinator Relationship, Mullerian mimicry, Batesian mimicry, Xanthopan morganii, Angraecum sesquipedale, Heliconius, Hymenopus coronatus, Papilio dardanus, Vachellia cornigera, Acacia cornigera, Pseudomyrmex ferruginea, Nepenthes bicalcarata, Colobopsis schmitzi, Camponotus schmitzi, Myrmecophyte, Domatia, Beltian Bodies, Extrafloral Nectaries, P R Ehrlich, P Raven, D H Janzen, E Bernays, M Graham, T Jermy, J N Thompson, C Darwin, A R Wallace, M A Rodriguez-Girones, L Santamaria, A L Llandres
http://www.gettyimages.com/detail/540976761
King Spider Orchid (Caladenia pectinata) being visited by male parasitic wasp pollinator which is attracted to the flower by faux female wasp pheromone, Australia
http://www.gettyimages.com/detail/540976675
King Spider Orchid (Caladenia pectinata) being visited by male parasitic wasp which is attracted to the flower by faux female wasp pheromone, western Australia
http://www.gettyimages.com/detail/540976787
King Spider Orchid (Caladenia pectinata) flower swarmed by male parasitic wasps who interpret the flower’s red lip as female wasp, western Australia
http://www.gettyimages.com/detail/540976759
Warty Hammer Orchid (Drakaea livida) flower has hinge mechanism to manipulate its pollinator into just the right position to receive its pollen, south of Perth, western Australia
http://www.gettyimages.com/detail/540976661
Zebra Orchid (Caladenia cairnsiana) flower being pollinated by male parasitic wasp which is attracted to the flower by faux female wasp pheromone, western Australia
http://www.gettyimages.com/detail/479670033
Orchid Bee (Euglossa sp) male visiting Orchid (Mormodes sp) flower
http://www.gettyimages.com/detail/475172281
Orchid Bee (Euglossa sp) male visiting Orchid (Mormodes sp) flower, Santa Rita, Panama
http://www.gettyimages.com/detail/540976735
Orchid Bee (Euglossa sp) male with orchid pollinia attached to its back, Barro Colorado Island, Panama
http://www.gettyimages.com/detail/540976795
Bucket Orchid (Coryanthes panamensis) flower attracting male Bee (Apidae) with sweet scent showing pollenia on its back, Gamboa, central Panama
http://www.gettyimages.com/detail/540976677
Orchid (Catasetum viridiflavum) with male orchid bee approaching, Barro Colorado Island, Panama
http://www.gettyimages.com/detail/540976747
Orchid (Masdevallia caloptera) flower with its pollinator Blue Bottle Fly (Calliphoridae) carrying yellow pollinium on its back, Finca Dracula Orchid Sanctuary, western Panama
http://www.gettyimages.com/detail/540976713
Bumblebee Orchid (Ophrys bombyliflora) flower and its male Longhorn Bee (Eucera sp) pollinator, Sardinia, Italy
http://www.gettyimages.com/detail/540976715
Bumblebee Orchid (Ophrys bombyliflora) flower and its male Longhorn Bee (Eucera sp) pollinator, Sardinia, Italy
http://www.gettyimages.com/detail/540976807
Brown Bee Orchid (Ophrys iricolor) flower mimics female bee with male solitary bee engaged in pseudocopulation, Sardinia, Italy
http://www.gettyimages.com/detail/475172159
Orchid (Himantoglossum robertianum) flowers with bee pollinator, Sardinia, Italy
http://www.gettyimages.com/detail/127475011
Olive Skipper (Pyrgus serratulae) butterfly on orchid, Hohe Tauern National Park, Austria
Embed from Getty Images
Brimstone butterfly (Gonepteryx rhamni) feeding on Fragrant Orchid, UK
http://www.gettyimages.com/detail/123804058
Bee pollinating a common spotted orchid (Dactylorhiza fuchsii). This process of pollination is called Entomophily, whereby pollen is inadvertently distributed by insects to other flowers. Photographed in Kent, UK.
http://www.gettyimages.com/detail/123758493
Common blue butterfly (Polyommatus icarus) feeding on a fragrant orchid (Gymnadenia conopsea). Photographed in June in Kent, UK.
Embed from Getty Images
Female common blue butterfly (Polyommatus icarus) resting on a southern marsh orchid (Dactylorhiza praetermissa), Dawlish Warren Nature Reserve, Devon, England
http://www.gettyimages.com/detail/135604800
Pair of marsh fritillary butterflies (Euphydrayas aurinia) resting on a heath orchid flower (Dactylorhiza fuchsii) in the countryside of Devonshire.
http://www.gettyimages.com/detail/135600799
Five-spot burnet moths (Zygaena trifolii) mating on Pyramidal orchid (Anacamptis pyramidalis) in Cornish countryside

Stylidium hortiorum with pollinator/robber. (Photo credit: Wikipedia)

Cycnoches haagii, Suriname. Pollinator: a bee from the Euglossini, also called orchid bees (Photo credit: Wikipedia)

Mormodes buccinator, Suriname. Pollinator: a bee from the Euglossini, also called orchid bees (Photo credit: Wikipedia)

Peristeria guttata, Suriname. Pollinator: a bee from the Euglossini, also called orchid bees (Photo credit: Wikipedia)
Related Sites and Articles
- Do Plants and Insects Coevolve? 🥀🐝🌺🦋 (queenslandorchid.wordpress.com)
- Do plants and pollinators coevolve? (standingoutinmyfield.wordpress.com)
- Flowers and insects: an ancient alliance (rcannon992.com)
- ‘Good Heavens what insect can suck it’- Charles Darwin, Angraecum sesquipedale and Xanthopan morganii praedicta (academia.edu) Joseph Arditti, John Elliott, Ian J. Kitching, and Lutz T. Wasserthal (2012). ‘Good Heavens what insect can suck it’- Charles Darwin, Angraecum sesquipedale and Xanthopan morganii praedicta. Botanical Journal of the Linnean Society, 169, 403–432.
- “Good Heavens, what insect can suck it…” (standingoutinmyfield.wordpress.com)
- “The Orchids Have Been A Splendid Sport” — An Alternative Look at Charles Darwin’s Contribution to Orchid Biology (academia.edu) Tim Wing Yam, Joseph Arditti, and Kenneth M. Cameron (2009). “The Orchids Have Been A Splendid Sport” — An Alternative Look at Charles Darwin’s Contribution to Orchid Biology. American Journal of Botany 96(12): 2128–2154.
- The Origin of Modern Biodiversity: Coevolution of Flowers and Insects (bioteaching.wordpress.com)
- Fine-tuned Bee-Flower Coevolutionary State Hidden within Multiple Pollination Interactions (nature.com) Akira Shimizu, Ikumi Dohzono, Masayoshi Nakaji, Derek A. Roff, Donald G. Miller III, Sara Osato, Takuya Yajima, Shûhei Niitsu, Nozomu Utsugi, Takashi Sugawara and Jin Yoshimura (2014). Fine-tuned Bee-Flower Coevolutionary State Hidden within Multiple Pollination Interactions. Scientific Reports 4, Article number: 3988. doi:10.1038/srep03988
- Coevolution (lifeofplant.blogspot.com.au)
- Interplay between insects and plants: dynamic and complex interactions that have coevolved over millions of years but act in milliseconds (jxb.oxfordjournals.org) Also available at http://jxb.oxfordjournals.org/content/66/2/455.full.pdf+html
- Did insect pollination cause increased seed plant diversity? (wolfweb.unr.edu) Root Gorelick (2001). Did insect pollination cause increased seed plant diversity? Biological Journal of the Linnean Society, 74: 407-427.
- Coevolution and Pollination (wnps.org)
- Interactions between Herbivorous Insects and Plant-Insect Coevolution (sites.biology.duke.edu)
- Coevolution (en.wikipedia.org)
- Mutualism (biology) (en.wikipedia.org)
- What is a mutualism? (standingoutinmyfield.wordpress.com)
- Coevolution of Mutualism Between Ants and Acacias in Central America. (jstor.org) Janzen D.H. (1966) Coevolution of Mutualism Between Ants and Acacias in Central America. Evolution 20: 249-275.
- Coevolution (britannica.com)
- Coevolution (bio.miami.edu)
- Lecture 9. Plant-Insect Interactions (ag.arizona.edu)
- Sweet, sweet coevolution (standingoutinmyfield.wordpress.com)
- Coevolution (pineapplesandwhales.wordpress.com)
- Geographic mosaic theory of coevolution (britannica.com)
- Orchid (britannica.com)
- Mimicry (britannica.com)
- The Geographic Mosaic of Coevolution (thompsonlab.sites.ucsc.edu)
- Geographic mosaic theory of coevolution (nature.com) Hoeksema, J. D. (2012) Geographic Mosaics of Coevolution. Nature Education Knowledge 3(10):19
- Coevolution: The Geographic Mosaic of Coevolutionary Arms Races (sciencedirect.com) John N Thompson. Coevolution: The Geographic Mosaic of Coevolutionary Arms Races. Current Biology, Volume 15, Issue 24, 24 December 2005, Pages R992–R994
- The Geographic Mosaic of Coevolution (scielo.org.ar)
- Plant-insect interactions: an evolutionary arms race between two distinct defense mechanisms (scielo.org.br)
- Research proposal: Predicting coevolutionary patterns on a global scale (spsasevo.wikidot.com)
- Evolution for beginners 2: coevolution (allyouneedisbiology.wordpress.com)
- Plants and animals can also live in marriage (allyouneedisbiology.wordpress.com)
- Orchids: different colours and shapes for everyone (allyouneedisbiology.wordpress.com)
- When Is It Coevolution? (bio.miami.edu) Janzen, D H (1980) When Is It Coevolution? Evolution 34: 611-612″. Also available from biologia.bio.br
- Resource Competition Triggers the Co-Evolution of Long Tongues and Deep Corolla Tubes (journals.plos.org)
- Book Review: “Pollination and Floral Ecology” by Pat Willmer (jeffollerton.wordpress.com)
- Nickolas M. Waser, Jeff. Ollerton, eds. 2006. Plant–Pollinator Interactions: From Specialization to Generalization. University of Chicago Press. Chicago, Illinois. (books.google.com.au)
- Book Review: “Plant–Pollinator Interactions: From Specialization to Generalization” by Carlos Martinez Del Rio (bioone.org) Also available in pdf.
- Book Review: “Plant–Pollinator Interactions: From Specialization to Generalization” by Paul D Rymer (researchgate.net)
- Book Review: “Plant–Pollinator Interactions: From Specialization to Generalization” by Peter Klinkhamer (ncbi.nlm.nih.gov)
- Pollinators and pollination – something for the weekend #9 (jeffollerton.wordpress.com)
- Fossil Focus: Arthropod–plant interactions (palaeontologyonline.com) Slater, B. J. 2014. Fossil Focus: Arthropod–plant interactions. Palaeontology Online, Volume 4, Article 5, 1-17.
- Orchid Mantis: Imitation and Disguise (queenslandorchid.wordpress.com)
- Secrets of the orchid mantis revealed – it doesn’t mimic an orchid after all (jdjgilbert.wordpress.com)
- Secrets of the orchid mantis revealed – it doesn’t mimic an orchid after all (theconversation.com)
- Pollinator Deception in the Orchid Mantis (jstor.org) O’Hanlon, J., Holwell, G., Herberstein, M., & Natural History Editor: Mark A. McPeek (2014). Pollinator Deception in the Orchid Mantis. The American Naturalist, 183(1), 126-132. doi:1. Retrieved from http://www.jstor.org/stable/10.1086/673858 doi:1
- Predatory pollinator deception: Does the orchid mantis resemble a model species (currentzoology.org) [Note: The link seems to be defunct.] J. C. O’Hanlon, G. I. Holwell, M. E. Herberstein (2014). Predatory pollinator deception: Does the orchid mantis resemble a model species. Current Zoology 60(1): 90–103.
- The evolution of imperfect mimicry in hoverflies (eprints.nottingham.ac.uk) Gilbert, Francis (2004). The evolution of imperfect mimicry in hoverflies. Insect Evolutionary Biology. CABI. (In Press)
- The Roles of Colour and Shape in Pollinator Deception in the Orchid Mantis Hymenopus coronatus (onlinelibrary.wiley.com)
- Orchid Mantis Perfectly Mimics Flowers In Order To Lure Prey [VIDEO] (isciencetimes.com)
- Mantis: las reinas del mimetismo (allyouneedisbiology.wordpress.com)
- Mimicry (en.wikipedia.org)
- Batesian mimicry (en.wikipedia.org)
- Müllerian mimicry (en.wikipedia.org)
- Aggressive mimicry (en.wikipedia.org)
- Wings of mimicry (desertspaces.wordpress.com)
- New paper: genetics of red pattern elements in Heliconius erato and Heliconius melpomene (heliconius.org)
- Butterfly Genetics Group: Martin Thompson (heliconius.zoo.cam.ac.uk)
- Darwin, C. R. 1862. On the various contrivances by which British and foreign orchids are fertilised by insects, and on the good effects of intercrossing. London: John Murray. (darwin-online.org.uk)
- Darwin, C. R. (1877). The various contrivances by which orchids are fertilised by insects. London: John Murray. 2d edition. (darwin-online.org.uk)
- Guest Blog: Darwin and the Lost Art of the Naturalist (movingsushi.com)
- Does science need more mavericks, or are they part of the problem? (aeon.co)
- Angraecum sesquipedale (en.wikipedia.org)
- Angraecum sesquipedale (Christmas star orchid) (kew.org)
- Darwin’s orchid (Angraecum sesquipedale) (arkive.org)
- Xanthopan (en.wikipedia.org)
- Moth tongues, orchids and Darwin – the predictive power of evolution (theguardian.com)
- Hawk-moth scale analysis and pollination specialization in the epilithic Malagasy endemic Aerangis ellisii (Reichenb. fil.) Schltr. (Orchidaceae) (onlinelibrary.wiley.com) Nilsson, L. A. and Rabakonandrianina, E. (1988). Hawk-moth scale analysis and pollination specialization in the epilithic Malagasy endemic Aerangis ellisii (Reichenb. fil.) Schltr. (Orchidaceae). Botanical Journal of the Linnean Society, 97: 49–61. doi:10.1111/j.1095-8339.1988.tb01686.x
- Exact tracking of pollen transfer and mating in plants (nature.com) Nilsson, L. A., Rabakonandrianina, E. and Pettersson, B. (1992). Exact tracking of pollen transfer and mating in plants. Nature 360, 666-668 (17 December 1992); doi:10.1038/360666a0
- The Scents and Smells of Orchids ❀👃✾ (queenslandorchid.wordpress.com)
- Evolving Digital Ecological Networks (journals.plos.org)
- The Genetic Basis of Pollinator Adaptation in a Sexually Deceptive Orchid (journals.plos.org)
- Deceptive Beauties – The World of Wild Orchids (blog.nationalgeographic.org)
- Trends in the pollination ecology of the Orchidaceae: evolution and systematics (researchgate.net) Raymond Louis Tremblay (1992). Trends in the pollination ecology of the Orchidaceae: evolution and systematics. Canadian Journal of Botany, 70(3): 642-650. https://doi.org/10.1139/b92-083
- Generalization in Pollination Systems, and Why it Matters (jstor.org) Nikolas M. Waser, Lars Chittka, Mary V. Price, Neal M. Williams and Jeff Ollerton (1996). Generalization in Pollination Systems, and Why it Matters. Ecology, 77(4), 1043-1060. doi:1. Retrieved from http://www.jstor.org/stable/2265575 doi:1. Also available from eebweb.arizona.edu
- Reconciling Ecological Processes with Phylogenetic Patterns: The Apparent Paradox of Plant-Pollinator Systems (jstor.org) Ollerton, Jeff (1996). Reconciling Ecological Processes with Phylogenetic Patterns: The Apparent Paradox of Plant-Pollinator Systems. Journal of Ecology, 84(5), 767-769.
- The explosion in orchids as houseplants: what does it tell us about how flowers evolve? (jeffollerton.wordpress.com)
- Variation in sexual reproduction in orchids and its evolutionary consequences: a spasmodic journey to diversification (academic.oup.com) Tremblay RL, Ackerman JD, Zimmerman JK, Calvo RN. (2005). Variation in sexual reproduction in orchids and its evolutionary consequences: a spasmodic journey to diversification. Biological Journal of the Linnean Society, 84, 1–54. Also available from researchgate.net
- Dispelling the myth that orchid species usually only have a single pollinator (jeffollerton.wordpress.com)
- The Pollination Ecology of an Assemblage of Grassland Asclepiads in South Africa (aob.oxfordjournals.org) Jeff Ollerton, Steven D. Johnson, Louise Cranmer and Sam Kellie (2003). The Pollination Ecology of an Assemblage of Grassland Asclepiads in South Africa. Ann Bot 92 (6): 807-834. doi: 10.1093/aob/mcg206
- Hot Spots, Cold Spots, and the Geographic Mosaic Theory of Coevolution. (ncbi.nlm.nih.gov) Gomulkiewicz R, Thompson JN, Holt RD, Nuismer SL, Hochberg ME (2003). Hot Spots, Cold Spots, and the Geographic Mosaic Theory of Coevolution. Ann Bot 92 (6): 807-834. doi: 10.1093/aob/mcg206
- Coevolution: Plant–Insect (els.net)
- Coevolution of angiosperms and insects (palaeo.gly.bris.ac.uk)
- Coevolution of Insects and (Flowering) Plants (biol.wwu.edu)
- Coevolution (sonoma.edu)
- Coevolution (evolution.berkeley.edu)
- Biodiversity and Plant-Animal Coevolution (press.princeton.edu)
- BIOL 2007 – COEVOLUTION (ucl.ac.uk)
- Natural Selection, Coevolution, and the Web of Life (jstor.org) Thompson, J. (2014). Natural Selection, Coevolution, and the Web of Life. The American Naturalist, 183(1), Iv-V. doi:1. Retrieved from http://www.jstor.org/stable/10.1086/674238 doi:1
- Evolution for the Community Ecologist (jstor.org) Callaway, R. (2006). Evolution for the Community Ecologist. BioScience, 56(3), 266-267. doi:1. Retrieved from http://www.jstor.org/stable/10.1641/0006-3568(2006)056[0266:eftce]2.0.co;2 doi:1
- Imperfect Batesian Mimicry and the Conspicuousness Costs of Mimetic Resemblance (jstor.org) Speed, M., Ruxton, G., Associate Editor: Franz J. Weissing, & Editor: Donald L. DeAngelis (2010). Imperfect Batesian Mimicry and the Conspicuousness Costs of Mimetic Resemblance. The American Naturalist, 176(1), E1-E14. doi:1. Retrieved from http://www.jstor.org/stable/10.1086/652990 doi:1
- Optimal Control and Cold War Dynamics between Plant and Herbivore (jstor.org) Low, C., Ellner, S., Holden, M., Associate Editor: William F. Morris, & Editor: Judith L. Bronstein (2013). Optimal Control and Cold War Dynamics between Plant and Herbivore. The American Naturalist, 182(2), E25-E39. doi:1. Retrieved from http://www.jstor.org/stable/10.1086/670810 doi:1
- Biotic Interactions, Rapid Evolution, and the Establishment of Introduced Species (jstor.org) Jones, E., Gomulkiewicz, R., Associate Editor: Diane S. Srivastava, & Editor: Mark A. McPeek (2012). Biotic Interactions, Rapid Evolution, and the Establishment of Introduced Species. The American Naturalist, 179(2), E28-E36. doi:1. Retrieved from http://www.jstor.org/stable/10.1086/663678 doi:1
- Source Populations Act as Coevolutionary Pacemakers in Experimental Selection Mosaics Containing Hotspots and Coldspots (jstor.org) Vogwill, T., Fenton, A., Buckling, A., Hochberg, M., Brockhurst, M., Associate Editor: Craig W. Benkman, & Editor: Michael C. Whitlock (2009). Source Populations Act as Coevolutionary Pacemakers in Experimental Selection Mosaics Containing Hotspots and Coldspots. The American Naturalist, 173(5), E171-E176. doi:1. Retrieved from http://www.jstor.org/stable/10.1086/597374 doi:1
- Multidimensional (Co)Evolutionary Stability (jstor.org) Débarre, F., Nuismer, S., Doebeli, M., Associate Editor: Maria R. Servedio, & Editor: Troy Day (2014). Multidimensional (Co)Evolutionary Stability. The American Naturalist, 184(2), 158-171. doi:1. Retrieved from http://www.jstor.org/stable/10.1086/677137 doi:1
- Comparative Dating of Attine Ant and Lepiotaceous Cultivar Phylogenies Reveals Coevolutionary Synchrony and Discord (jstor.org) Mikheyev, A., Mueller, U., Abbot, P., Associate Editor: Jukka Jokela, & Editor: Ruth G. Shaw (2010). Comparative Dating of Attine Ant and Lepiotaceous Cultivar Phylogenies Reveals Coevolutionary Synchrony and Discord. The American Naturalist, 175(6), E126-E133. doi:1. Retrieved from http://www.jstor.org/stable/10.1086/652472 doi:1
- Inverse-Gene-for-Gene Infection Genetics and Coevolutionary Dynamics (jstor.org) Fenton, A., Antonovics, J., Brockhurst, M., Associate Editor: Greg Dwyer, & Editor: Mark A. McPeek (2009). Inverse-Gene-for-Gene Infection Genetics and Coevolutionary Dynamics. The American Naturalist, 174(6), E230-E242. doi:1. Retrieved from http://www.jstor.org/stable/10.1086/645087 doi:1
- Herbivory and Defensive Characteristics of Tree Species in a Lowland Tropical Forest (jstor.org) Coley, P. (1983). Herbivory and Defensive Characteristics of Tree Species in a Lowland Tropical Forest. Ecological Monographs, 53(2), 209-229. doi:1. Retrieved from http://www.jstor.org/stable/1942495 doi:1
- Macroevolution and the biological diversity of plants and herbivores (pnas.org) Douglas J. Futuymaa and Anurag A. Agrawal (27 October 2009). PNAS 2009 106 (43) 18054-18061, doi:10.1073/pnas.0904106106. Plant and Insect Biodiversity Special Feature (free online). Retrieved from http://www.pnas.org/content/106/43/18054.full
- Diversification and coevolution in brood pollination mutualisms: Windows into the role of biotic interactions in generating biological diversity (amjbot.org) David H. Hembry and David M. Althoff (2016). Am. J. Bot. October 2016 vol. 103 no. 10 1783-1792. Retrieved from http://www.amjbot.org/content/103/10/1783.full
- Orchid Sexual Deceit Provokes Ejaculation (jstor.org) Gaskett, A., Winnick, C., Herberstein, M., & Natural History Editor: Henry M. Wilbur (2008). Orchid Sexual Deceit Provokes Ejaculation. The American Naturalist, 171(6), E206-E212. doi:1. Retrieved from http://www.jstor.org/stable/10.1086/587532 doi:1
- Pollination and evolution of the genus Mystacidium (Orchidaceae) (researchspace.ukzn.ac.za)
- Mystacidium capense: A Dainty Orchid from South Africa 🌍✾ (queenslandorchid.wordpress.com)
- Prosthechea cochleata: An Upside-Down, Dangly and Charming Orchid ✾ (queenslandorchid.wordpress.com)
- The Living, Breathing World of Borneo’s Carnivorous Pitcher Plants (proof.nationalgeographic.com)
- The Beauty of Deceit (phenomena.nationalgeographic.com)
- Cool Bugs #9 – Acacia ants (bioblog.biotunes.org)
- Beetle-Pollination ─ Cantharophlly ─ in Amorpbopballus bobenackeri (Araceae) (aroid.org)
- Jonathan Drori: The beautiful tricks of flowers (ted.com)
- Co-evolution with dinosaurs? (palaeo.gly.bris.ac.uk)
- The Vegetarian Spider (standingoutinmyfield.wordpress.com)
- Yucca Moths (standingoutinmyfield.wordpress.com)
- Tough Questions (dailypost.wordpress.com)
- The Poetry of List-Making (dailypost.wordpress.com)
- Mixing Media (dailypost.wordpress.com)
- Symbiosis (dailypost.wordpress.com)
- 100 Best Things to do in South Africa (jenreviews.com)














Pages: 1 2

[…] Do Plants and Insects Coevolve? (soundeagle.wordpress.com) […]
LikeLiked by 3 people
[…] Do Plants and Insects Coevolve? (soundeagle.wordpress.com) […]
LikeLiked by 1 person
[…] Do Plants and Insects Coevolve? (soundeagle.wordpress.com) […]
LikeLiked by 1 person
Co evolution and symbiotic relationships are very common aspects of biology. A very fascinating area of study.
LikeLiked by 10 people
Moreover, there is another version of this post published at https://queenslandorchid.wordpress.com/2016/08/17/do-plants-and-insects-coevolve/ where you will find the discussions at the comment section very engaging, and where there are seven superb videos available for viewing.
LikeLiked by 2 people
Hi Sound Eagle,
You liked a comment I made on the Steve Says blog. I wanted to come over to say hi and introduce myself. Hi, I am Janice.
This seems like a great site for science lovers. My daughter is into science. I teach history and overlap with science often.
Nice to meet you.
Janice
LikeLiked by 7 people
LikeLiked by 3 people
There are so many examples in nature of these symbiotic (I think that is the word) relationships that you can easily see how much by design it all is. We have a swan plant in the garden here in Auckland which, as I understand it, is the only plant monarch butterfly caterpillars eat, perhaps another example…although I am not sure what benefit the plant gets from this.
Also, going a little further than insects, I look at the Tui (NZ bird) drinking nectar from a flax plant and think how the beak is perfectly shaped and sized to fit into the flower buds. There are loads of examples in nature I suppose.
But where does mankind fit in to this…well…not sure we do…we are kind of embarrassing where ecosystems are concerned.
LikeLiked by 7 people
Thank you for your interest in our article, Graham. It is quite common for insects to eat only a narrow range of plant species. Monarch larvae specialise in plants of the family Asclepiadaceae (commonly called milkweeds). Not all ecological relationships are mutually beneficial; probably the plant does not gain anything from this relationship, except perhaps some help from the adult butterflies in pollination.
Clear examples of coevolutionary relation ships between humans and other organisms are hard to find. One possible example may be with some strains of gut micro-organisms. Certainly, some of these seem to have important positive effects on human health, but I don’t know to what extent they have become adapted specifically to the environment of the human digestive tract.
LikeLiked by 8 people
In symbiogenesis (or endosymbiotic theory), the evolution of eukaryotic cell in animals, plants and fungi could be considered as a case of coevolution to the extent that it began as a symbiotic community of prokaryotic cells or organisms. For example, inside plant cells are DNA-bearing organelles like the mitochondria and the chloroplasts, both descended and coevolved from ancient symbiotic oxygen-breathing proteobacteria and cyanobacteria respectively, and both were endosymbiosed by an ancestral archaean prokaryote, which (co)evolved into the cellular membrane.
Some scientists, especially those in the interdisciplinary fields of anthrozoology and zooanthropology, may be more inclined to entertain or conclude that the ongoing domestication processes involving humans and animals have been significant coevolutionary forces at work, especially with respect to the development and evolution of human civilization, and to the phenotypic variation and diversity of domesticated animals and pets. By the same token, plants such as wheat, rice and other staple crops and cultivated grasses (including the banana) as well as some hybridized or genetically modified ornamentals and food plants have been dramatically altered even to the point of being sterile or losing their self-seeding ability, their survival relying solely on being cultivated by humans. On the whole, the domestication of animals and plants has caused a rapid shift in the evolution, ecology and demography of both humans and numerous species of animals and plants. Like their human counterpart, these domesticated species have become some of the most abundant and widespread organisms on Earth.
LikeLiked by 3 people
That’s a very fascinating and detailed post. I’ll have to reread this more closely later.
LikeLiked by 6 people
May you enjoy springtime in Hong Kong excavating even more fun (and surprising) facts as the weather warms up!
LikeLiked by 3 people
The most pertinent and/or awe-inspiring fun (and surprising) facts might eventually be incorporated into the body of this post.
LikeLiked by 1 person
Thank you for sharing this brilliant article. And thanks for the links as well (I read most of the articles which you have linked). I have learned a lot.
I have been familiar with the concept of coevolution for quite some time but I didn’t give much thought about it. After reading this article and the linked articles, it made me ponder about this topic more.
Of course, it’s difficult to prove coevolution but I think that they did occur at some point, we may never know when. Based on the examples that this article has enumerated, it’s difficult to not give coevolution some consideration.
These links may interest you:
http://proof.nationalgeographic.com/2015/01/28/the-living-breathing-world-of-borneos-carnivorous-pitcher-plants/
http://bioblog.biotunes.org/bioblog/2007/10/02/cool-bug-9-acacia-ants/
Click to access 0121411.pdf
Regarding specialization, I can relate to that since this is also the case in mathematics. Even specialized fields like Algebraic Topology has many subfields. We may never see the likes of Archimedes and Euler. Perhaps Henri Poincaré and Hilbert were some of the last people who have a great understanding of the different branches of mathematics. Clifford Pickover speculated that the most knowledgeable mathematicians alive today may only know about 5% of the entire corpus of mathematics, though I suspect that it’s a lot less.
Having so many types of “-ologists” out there cause most people to not care about their research that much because they are too specialized for the “outsiders” to comprehend. As Henry Ward observed, “An expert is one who knows so much about so little that he neither can be contradicted nor is worth contradicting.”
LikeLiked by 7 people
Hello Edmark! It has been a while since we interacted here regarding the “fun facts (and figures)” of the coevolution of plants and insects. SoundEagle would like to take this opportunity to inform you that the expansive Foreword by SoundEagle has been further improved and extended.
You can probably still remember SoundEagle mentioning to you in this comment that “[t]he most pertinent and/or awe-inspiring fun (and surprising) facts might eventually be incorporated into the body of this post.” Your feedback is indeed a testament to your engaging and productive nature, as can be seen in your avid response to SoundEagle welcoming you “to include within your forthcoming feedback on this post many fun (and surprising) facts about orchids, (co)evolution and/or any other subject matters discussed in this post.”
Indeed, your excellent and multifaceted feedback, which contains several informative weblinks, has brought positive outcome and further impetus to this post and its twin. Upon visiting the corresponding webpages and consulting additional sources, SoundEagle learnt that “myrmecophytes”[11] are indeed valid examples of coevolution, and therefore prompted Dr Craig Eisemann to add a new section, which he completed and titled “Ants and Plants”. Please be informed that both Dr Eisemann and SoundEagle appreciate your feedback and would like to express our gratitude towards you here.
In addition, your matching SoundEagle’s comparison of Charles Darwin and modern-day academics/researchers specialized in natural science (or science in general) with your parallel comparison of generalists such as Archimedes and Euler against modern-day highly specialized mathematicians, highlights some of the most intractable predicaments of conducting research and being researchers, many of which can readily lead to even more serious ramifications in the future, unless certain long-term solutions can be found or engendered through artificial intelligence, neural networks and expert systems.
Undoubtedly, our interactions in the comment section of “Do Plants and Insects Coevolve? 🥀🐝🌺🦋” demonstrate that there are plenty of common interests and intellectual momenta here to resonate with the caveats and concerns discussed in certain parts of the Foreword by SoundEagle as well as your esteemed feedback and SoundEagle’s first, second and third comments on set theory in your post entitled “101 Mathematical Trivia”. Therefore, SoundEagle would like to augment all of our comments, discussions and contributions with the following articles, whilst acknowledging some of the most influential aspects, issues and relationships within and between arts, sciences and mathematics:
LikeLiked by 2 people
I have seen few blog sites that have placed this much time and effort into creating an experience that reaches the depth of real sensory stimulation. I commend ALL your effort. Wonderful post!
LikeLiked by 8 people
Hello Dr Jonathan! As the festive season is drawing to a close and February is just around the corner, SoundEagle hopes that you have had a wonderful time celebrating the dawn of 2018 whilst having a great holiday with your family and friends.
Thank you for your visit and compliment, and for being observant about, and receptive towards, the presentational and experiential aspects of the post, beyond what its diverse and analytical contents convey about the coevolution of plants and insects 🥀🐝🌺🦋. Hence, SoundEagle has good reasons to be truly delighted by your regarding the post to be wonderful.
Also comparable in quality, expanse and “the depth of real sensory stimulation” is the most recent post, aptly named The Quotation Fallacy “💬”, which can firmly constitute excellent food for thought as well as a splendid guide for living a more examined life. May you find the post as enjoyable as it is edifying!
LikeLiked by 2 people
[…] Do Plants and Insects Coevolve? 🥀🐝🌺🦋 (soundeagle.wordpress.com) […]
LikeLike
[…] Do Plants and Insects Coevolve? 🥀🐝🌺🦋 (soundeagle.wordpress.com) […]
LikeLike
Excellent. Thank you.
LikeLiked by 5 people
Hello Gary,
Sorry for the very delayed reply. We are glad to have produced a post that you found useful. I have recently looked at your excellent website. You may be interested in some forthcoming posts, to do with environmental issues, evolutionary effects on human behaviours and human responses to the natural world, as well as various posts by SoundEagle🦅 on diverse topics that are already up on this site.
LikeLiked by 1 person
Thanks for teaching me!
LikeLiked by 5 people
Hello Peter,
My apologies for the very late response. We are pleased to have produced a post that was of interest to you. I have found that the more I learn about biological organisms and processes, the more I come to appreciate the beauty of their diversity and intricate adaptations. Please consider looking at other posts on this site, and also some forthcoming ones to do with environmental issues, human behaviours and human psychological responses to Nature.
LikeLiked by 4 people
God made them all – all organisms – in their simplicities and complexities. God is behind them all – all happenings – in their endothermicities and exothermicities.
LikeLike
A comprehensive demonstration that diversity is a source of strength, not weakness, a proven fact which certain modern political and religious creeds seek to sidestep!
LikeLiked by 6 people
Hello Dave
Diversity is certainly a source of resilience in ecosystems, and it seems quite reasonable that a similar principle also applies to other systems, including human societies. We hope you will also find some interest in the quite diverse posts by SoundEagle🦅 on this site, as well as some forthcoming ones on topics such as human nature, human appreciation of the natural world and sustainability and conservation. Apologies for the very late response.
LikeLiked by 2 people
Hi, and sorry for my reply delay! I will certainly return to your site as science, a key to future understanding, underpins your work.
LikeLiked by 2 people
Thank you, Dave; I hope we will be able to meet your expectations in forthcoming posts. Looking forward to your future comments.
LikeLiked by 2 people
Thank you Dave, I hope we can meet your expectations in future posts.Looking forward to your comments in future
LikeLiked by 2 people
Beautiful examples of something I would not think many people would disagree; indeed, it would be strange, if plants and insects would not co-evolve? i wonder if despite all these arguments, the phylogeny was done on these orchids? DNA sequence can reveal if there is an inter-species relationship or not. if yes, the role of insects is a more favorable factor. Continuing this thought, if flower preparation can be sequenced and the DNA of the same insect can be detected in several different species? Pure speculation of a potential experiment:))
another comment if on Wiki definition of abiotic factors that cannot be co-evolved since they are not living. Somewhere i read ( sorry cannot provide a ref) about one study of how animals and plants affect abiotic factors and those affecting them back- I do think this have a right to defined as co-evolvolution. but this indeed might spark arguments ( especially from evolutionary biologists:))
LikeLiked by 3 people
Hello Anastasia, and thank you for taking the time to read and comment on this article. Soundeagle has just alerted me to your comment and also wishes to convey his greetings.
I agree that it could reasonably be expected that organisms engaged in a close and mutually important ecological relationship would affect each other’s evolution. However, producing definite proof that this has occurred in particular cases can sometimes be difficult.
As regards your second question, although I am not sure that I fully understand your proposed experiment and admittedly speaking as a non-molecular biologist, I cannot see how DNA sequencing could shed much additional light on whether a coevolutionary relationship with another organism has developed. Related species such as plants or insects of a particular family or genus, whilst sharing much of their genetic material because of their phylogenetic relationship, may differ genetically from each other for a variety of reasons besides the development of coevolutionary relationships by some of their members, and the genetic basis of many of the characters selected in the development of such relationships may be complex and not understood. Even where the genetic basis of a character that is suspected to have evolved as a result of a coevolutionary relationship is known, its presence in the genome of an organism probably does not add any information than could be inferred from the presence of the character itself.
With regard to you third question, while it is certainly true that organisms are influenced in their evolution by abiotic factors and that many organisms can in turn affect abiotic factors (such as the composition of soils and of the atmosphere), the term evolution as used here is normally considered to apply only to living things, resulting as it does from natural selection operating ultimately on their genome. Any effects of organisms on abiotic components of the environment seem to constitute a fundamentally different process.
Once again, I am grateful for your interest, and hope that you will also read future posts on this site.
LikeLiked by 6 people
Hello Anastasia! Your comment is quite excellent indeed. It is heartening that you have given so much thought on the many aspects of coevolution discussed in this post, even mentioning DNA sequencing and consulting Wikipedia on the definition of abiotic factors. Your inquisitiveness and engaging attitude are to be highly commended, as is your effort in reading this protracted and multifaceted post.
SoundEagle wonders what your favourite plants and insects are. Do you grow some plants and keep certain insects?
Given the time of the year, here is a very special post with somewhat belated 🎊 Season Greetings from SoundEagle: Merry Christmas, Happy New Year and Joyful Holiday 🎄🎅⛄ to you and your family!
LikeLiked by 2 people
Hello SoundEagle,
Thank you so much for your reply . I personally do not keep any insects. Regarding plants, years ago I owned a cactus collection. It was amazing to try to cultivate them from seeds- I guess, that was as far as I went with it. Moving to a different country made it impossible to carry plants with me- so it went to friends…
I found the post super interesting- as many other readers. Especially the way you explain the phenomenon of co-evolution. To be honest, I am very happy that see lots of comments for your post from people of different backgrounds.
LikeLiked by 4 people
Just would like to add a little note about co-evolution. These days it becomes a common wisdom to take probiotics for a healthy digestion. So one of the pints is the co- evolution of microbiome and the organism. And we know, it does change now, when many of us eat sterile food and leave is cleaner conditions but in highly populated areas.
On another note, I wonder about your background? Are you a scientist?
LikeLiked by 4 people
Possible coevolutionary relationships between animals and their symbiotic microorganisms are an interesting topic for further research, especially in view of accumulating evidence that the composition of the gut microflora in particular has significant effects on human health.
Although not a professional scientist in the traditional sense as a result of going against the grain to adopt or create artful, novel, multidisciplinary and unorthodox approaches to researching, writing and publishing, SoundEagle nevertheless has a strong interest in many scientific questions and endeavours. In addition, SoundEagle has informed me that more scientists can and should be much better informed by the philosophy of science, the history of science, and the sociology of science.
On the other hand, I am a (rather prematurely) retired biologist, trained particularly in Entomology, but having interests in a wide variety of other subjects.
Please stay tuned for three additional posts to appear on this site fairly soon.
Best wishes, and thank you again for your interest.
LikeLiked by 5 people
You have not only great insight into problem but also beautifully written posts! I totally admire your ( both of you) writing. A talent plus education. Bravo!
LikeLiked by 3 people
Hi Anastasia! How have you been since the dawn of February?
Thank you for your keen interest in, and kind compliment on, our co-authored post here. As you can see, the numerous interrelationships and interspecies dynamics uncovered by certain scientists in the ongoing coevolution of plants and insects 🥀🐝🌺🦋 are far more intricate and manifold than most people have realised. What a multifarious affair coevolution can be!
Like you, those who are highly perceptive and very thorough with their reading of Do Plants and Insects Coevolve? 🥀🐝🌺🦋 can indeed become far more appreciative of the complexities incurred in answering such a question, which can significantly entail and benefit from going beyond the straightforward approach of presenting a series of major research findings and milestones through conducting a substantial literature review, insofar as the style, content and delivery of this post have demonstrated such entailments and benefits to a considerable degree via interactive, visually vibrant and intellectually engaging discussions in both the post and the comment section.
Please be invited to reread Dr Craig Eisemann’s amended and extended comment to you dated 30 January 2018 at 8:16 AM.
You are also cordially invited to be our esteemed reader of the following posts, chosen especially for you:
Given that St Valentine’s day is just around the corner, may you enjoy visiting 🦅 SoundEagle in Love and Dove, Art and Heart on Valentine’s Day with Gifts 🎁💝🕊💌💘!
LikeLiked by 2 people
Thank you so much! I will be more than happy to read it!!
LikeLiked by 4 people
Hello Anastasia! We have not heard from you and hope that life is treating you well in February, during which the big festival known as Lunar New Year is being celebrated all over the world for 16 days, as explicated in the encyclopaedic post 🦅 SoundEagle in Chinese New Year Celebration, Spring Festival, Lion Dance, Traditional Culture and Architecture 🏮🎋🦁⛩.
Given your academic interests in evolutionary biology, botany, biochemistry and genetics, the following posts published by SoundEagle🦅 at ✿❀ Queensland Orchid International ❀✿ are recommended to you:
LikeLiked by 2 people
Hello Anastasia
Thank you for your kind words. You may be interested to know that Soundeagle, who has considerable expertise in fields including Computer Science and Social Sciences, has a linked-in profile which can be viewed at
https://www.linkedin.com/in/soundeagle
I hope we can maintain or improve our standard in future posts!
Best regards
LikeLiked by 3 people
Hello Graig Eisemann and Sound Eagle! I am very happy to connect with you on LinkedIn ( I sent the invitation to the LinkedIn profile you mentioned in your post). Your point that there is a great need for scientific philosophy for scientists is extremely important. As a person who is busy with experiments and have little time to think more than my area of expertise ( and that is not very successful either), I feel that unbiased view on where we move and how this integrates with other aspects of our activities is a must! Have a nice week!
LikeLiked by 2 people
Hi Sound Eagle,
Look at how much time you put into all this. Quite a compilation! My husband watches many animal shows and my daughter loves science.
As far as I go, your eagle theme resonated with me. Once, in Alaska, I saw a bald eagle. I’d like to go to Alaska to see bears but I teach and they aren’t visible when I can go.
You liked a comment I made on Sam’s blog. I wanted to come by to say hi and introduce myself.
Janice
LikeLiked by 5 people
Hello Janice
Apologies for the very late reply. We are happy that you appreciated our efforts!. You may also be interested in some other posts on this site, including 🦅 SoundEagle in Debating Animal Artistry and Musicality 🎵🐕🎶🐒🎹🐘🖼🐬🎨. There should also be some new posts to do with evolution, Nature and conservation in the future.
LikeLiked by 2 people
I’m thoroughly impressed by your work here. Please pass my appreciation onto SoundEagle. WE have ospreys and golden eagles here in Scotland who send their greetings
LikeLiked by 6 people
Soundeagle and I are grateful for your kind comments. Please convey our best wishes to the Scottish raptors.
LikeLiked by 2 people
Wowee, what a blog! Very comprehensive. Insects and plants are so tight. As a silly thought-experiment, if insects evolved into the sentient organisms that we became, I can only imagine how influential plants would be in their art and culture, given their primordial co-relationship.
LikeLiked by 4 people
Hello Jesse
Thank you for reading and commenting on our post. It is interesting to consider what influences the biology of a hypothetical intelligent species might have on its culture. Edward O. Wilson once performed a thought experiment of this kind with termites, concluding that a termite species with human-like intelligence would, among other cultural values, abhor personal reproduction (except for the reproductive caste), embrace cannibalism of the aged and infirm and make a sacrament of the exchange and consumption of faeces.
However, it seems unlikely that an insect could attain a level of cognitive function similar to that of humans, at least without radical changes to its body plan and physiology. Humans have perhaps five orders of magnitude more brain neurons than even such a behaviourally complex insect as a honeybee. Miniaturization of neurons and increasing organizational efficiency of brain structures would have finite limits; the evolution of intelligence comparable to that of humans would therefore require a great increase in the size of the central nervous system and hence of the whole insect. This would most likely push the insect into a size range where its body plan would be much less efficient than that of similarly-sized vertebrates, so that inability to compete with these would probably terminate evolution in this direction well before human-like cognitive capacities were reached.
Nevertheless, the theoretical possibilities for the culture and cultural products of a cognitively advanced animal with a biology radically different from that of humans are fascinating to consider.
I hope you may be interested in some forthcoming posts on this site, the first to do with some human mental proclivities and their current effects on the biosphere.
Warmest regards and best wishes
Craig
LikeLiked by 4 people
Perhaps you could indeed imagine yourself as one of the “insects evolved into the sentient organisms that we became,” and then “imagine how influential plants would be in their art and culture, given their primordial co-relationship.” Looking forward to hearing from you and seeing your artworks in collaboration with SoundEagle🦅! Happy mid-September to you, Jesse!
LikeLike
Interesting thoughts. Mother nature never ceases to amaze. We humans need to take care!
LikeLiked by 3 people
We are pleased that our post inspired your interest. You may also like some other posts on this site, including 🦅 SoundEagle in Debating Animal Artistry and Musicality 🎵🐕🎶🐒🎹🐘🖼🐬🎨, and others, relating to evolution and environmental conservation, still to appear.
LikeLiked by 1 person
[…] via Do Plants and Insects Coevolve? 🥀🐝🌺🦋 […]
LikeLiked by 1 person
WOW! One of the most insightful and informative posts I have read to date. I love nature and we all coexist, like it or not. Thanks so much for sharing!
LikeLiked by 4 people
We are happy that you found our post to be of interest. The relationship between insects and plants has long been an interest of mine, and the study of their mutual adaptations has long helped to stimulate my fascination with living nature. You may also like to see other posts on this website, for instance “🦅 SoundEagle in Debating Animal Artistry and Musicality 🎵🐕🎶🐒🎹🐘🖼🐬🎨“.
LikeLiked by 1 person
Reblogged this on By the Mighty Mumford and commented:
IT IS TRUE–PLANTS AND ANIMALS EVOLVE OR ARE CREATED—TO CHANGE TO MAXIMUMLY INTERACT WITH THEIR ENVIRONMENT, OTHER ORGANISMS…AND RESPOND TO MANKIND’S FORCED MANIPULATION FOR HIS AND HER OWN PURPOSES. INDEED ALL ORGANISMS DO INTERACT IN ONE WAY OR OTHERS! SCHOLARLY AND DEFINITIVE WELL-DOCUMENTED RESEARCH. AND SPECIES DON’T CHANGE INTO OTHER SPECIES—BUT AS HERE, THEY ARE VERY ADJUSTABLE (OR NOT!). 🙂
LikeLiked by 4 people
A very comprehensive and fascinating affirmative answer to the question posed!
LikeLiked by 3 people
Please accept our belated thanks for your attention to our post. Much in evolutionary biology is of necessity rather speculative, but in the case of evoluionary interactions between these two groups of fascinating organisms, there seems to be quite strong evidence that coevolutionary processes have occurred.
You may also be interested in other posts by SoundEagle🦅 on this site, including 🦅 SoundEagle in Debating Animal Artistry and Musicality 🎵🐕🎶🐒🎹🐘🖼🐬🎨.
LikeLiked by 2 people
Dear friend,
I have nominated you for the “Mystery Blogger Award” – congratulation, my friend 🙂
For more details:
https://didisvgp.wordpress.com/2018/07/28/thank-you-for-the-mystery-blogger-award/
All good wishes
Didi
LikeLiked by 4 people
Your own response to the award, as seen at your recent post Thank you for the Mystery Blogger Award, is both amiable and amicable. You are now cordially invited to experience a special kind of response and writing from SoundEagle🦅 at this particular post:
LikeLiked by 2 people
Intriguingly fascinating art and thoughts. It’s different and I thoroughly enjoy unique writings- especially nature. Marvelous, simply marvelous!
LikeLiked by 4 people
Thank you, Mr Lance Sheridan, for your visit and compliment. You are receiving a very belated reply as it has been a very busy time. Happy mid-September to you!
Given your penchant for writing poems, SoundEagle🦅 would like to present you with an opportunity to write a poem to be featured here in this post. Assuming that you have indeed read the entire post, you would have noticed the featured poem at Co-Existence, a Poem by Cheryl KP. Stylistically and lengthwise, you are free to experiment. Your poem can be submitted via ✉ Email or via the Comments page.
In addition, you are also welcome to submit one or more poems based on your reading and appreciation of some or all of the following posts:
LikeLiked by 2 people
A pleasure! Your’s is an excellent site. I’m very grateful for your response!
LikeLiked by 2 people
A delightful encyclopedia of knowledge that fed my love of plants, Bees, Butterflies and Orchids.. Remarkable knowledge.. ❤
LikeLiked by 2 people
Dear Sue,
Belated thanks for your appreciation of our post. I have always found that an evolutionary perspective on living things tends to enhance my appreciation of their wonderful diversity, complexity and compatibility with their living and non-living environment. I hope you will be similarly gratified by other posts on this site, including some planned future ones.
LikeLiked by 2 people
You are welcome, and your blog is a mind of information. 🙂
LikeLiked by 2 people
This is a fascinating article, which I admit took me two days to read. It’s so densely packed with information!
The fact that Darwin predicted the existence of the Morgan’s Sphinx Moth forty years before it was discovered is poetic justice if I ever saw it. It begs the question, why did the flower’s morphology become so extreme? Surely if the adaptation were to exclude some non-beneficial insect, why didn’t its “nectariferous spurs” stop at 9 or 10 inches? Why did they have to go all the way to 11.5 inches?
I read through some of the papers you linked to but it’s tricky when you have to look up the definition of every second word haha! Although I didn’t find my answers, I’m grateful that you sent me on such an illuminating adventure. Articles like this open one’s eyes to the world. Thanks.
LikeLiked by 3 people
My apologies for the late reply. Thank you for making the effort to read and understand this article in such detail; I’m sure not everyone would do this! In answer to your question, it is sometimes difficult to deduce why biological structures take the exact form that they do. It may have to do with the fact that natural selection must work with whatever random mutations turn up, even if they are not optimal in terms of function or efficient use of resources. Alternatively, I suppose it is possible that another co-adapted orchid-hawkmoth association, involving different species, may once have occupied the middle ground (in terms of corolla tube and haustellum length) between the one discussed here and shorter, more usual lengths, but that these two hypothetical species became extinct relatively recently, not allowing enough time for the relevant structures in A. sesquipedale and X. morganii to shorten to a more economical length.
Greetings and best wishes for the new year to both of you
LikeLiked by 2 people
Hello Dear webmasters and readers;
Co-evolution is larger than most people guess;
It concerns all things on earth, and also all material in the universe;
The whole universe is ONE PROGRAMMED MACHINE running to a pre-defined goal;
and in each step during this course, human can observe new phenomenas,
just like in a train we can see so many different landscapes between each differents stations…
So the important for human to be in harmony with the universe is to observe objectively…
And good travel for you all ; Dear webmasters and reader…
LikeLiked by 3 people
Thank you for reading our post. It would be interesting to consider the evidence that the universe is evolving according to a pre-determined plan, rather than as a result of an unguided interplay between “chance and necessity”.
LikeLiked by 2 people
It is a pleasant fancy that humans are evolving to a greater fondness for cute animals and animals are evolving to look cuter to humans!
LikeLiked by 2 people
The recognition of “cuteness” probably evolved because it encouraged the protection and nurturance of offspring, and it is interesting that it can often cross species barriers, especially among mammals. In view of the threat of human-caused extinction that hangs over an increasing proportion of species, it may indeed be advantageous for them to appear “cute” to humans, as this may increase their chances of survival, both as individuals and as species!
LikeLiked by 2 people
Thank you for this very educational post! You obviously put a great deal of work and research into writing this. I can appreciate that! I think my brain may be bleeding now though…
LikeLiked by 2 people
We are grateful that you have read and appreciated our work! It must contain a lot of new information for anyone not already familiar with this field. Both of us learned a good deal in preparing this post, and are pleased if our readers can also learn something from it, even though it may cost a bit of effort! Best wishes, and hoping that you will also read some of our future posts.
LikeLiked by 2 people
Thank you. The post does contain much information that I was previously unfamiliar with. It is refreshing to find an educational blog about this subjects and I look forward to reading (slowly & with english dictionary at my table) your future posts!
LikeLiked by 3 people
Thank you, Mliae, for your visit, comments and compliment, as well as for recognizing the educational value of this special post. Whilst you passively wait for future posts to be published, you are very welcome to actively visit other educational posts to whet your intellectual appetite. Moreover, if your web browser can run Adobe Flash Player properly as it loads the contents of this website, then you will also be able to play games, solve puzzles and/or see more animations, which may indeed counteract over-exertion and stop your brain from bleeding. Happy February and Happy Exploring those posts!

Yours sincerely,
ჱܓSoundEagle🦅
LikeLike
I do not have time to do this lengthy article justice at the moment. In fact, I,d given up reading within the areas of academics or bordering upon it awhile ago, with the occasional exception of philosophy. But nature, genetics, evolution, Darwinism, and Neo-Darwinism (biology as ideology) are all areas I once was quite taken by. I noted too that you veered off into problems with the current structure of academics midway, and I too read B.V.’s article about why he departed the fold. (BV and I also had some dialog awhile back & he looked at one of my longer essays.) So, I will try and make time to get back to this post for a deeper commentary.
But meanwhile, I will mention that just last week I was watching one of David Attenborough’s wonderful video essays concerning plant/animal cooperations, and I was struck with an interesting intuition. The species involved were a kind of flower with deeply recessed nectar (I believe a red helliconia) and a specific humminbird with a purposed long curved beak. The relationship between these two were such that the bird fed exclusively upon that flower’s nectar while the flower’s relied exclusively upon that bird’s pollination techniques. An interesting all-your-eggs-in-one-basket evolutionary sidepool. The intuition, which I have not had time to develop and articulate better, had to do with the spiritual cooperation between the generating forces (the “ideas”) behind these two disparate species… how they must indeed have a singular and peculiar spiritual relationship in the world behind the physical. (This is fairly typical for me since I do not limit myself to a materialistic, i.e. exclusively physicalist worldview). Plants physicalized on earth first, before animals. So the external evolutionary story, which restricts itself to the purely physical, cannot explicate the entire story here. There is something to understand from a different angle. I leave it there for now. Hopefully I can return.
LikeLiked by 2 people
Hello Rob,
Thank you for your attention and thoughtful comments. For my part, I continue to be confident that explanations for biological processes can be found wholly within the physical realm. For example, I do not think that the appearance of two different groups of organisms at different times presents any difficulties for the subsequent development of coevolutionary relationships among some of their members – certain species of the second group to appear may develop ecological relationships with existing members of the first, influencing the further evolution of these and having their own evolutionary development affected by the interaction: it is not necessary for the relationships to begin upon the first appearance of both groups. Further, attempts to model the evolution of long sucking organs in various groups of animals and the corresponding evolution of long nectar-bearing corolla spurs in the flowers from which they obtain nectar have suggested that that this process is explicable in terms of conventional natural selection theory (please see my brief discussion of this and the references cited in this post).
It seems that your positing of the existence of non-material precursors of organisms in “the world beyond the physical” (and potential interactions among these) bears some relation to the Platonic theory of Forms. When you can find the time, you may like to elaborate upon these ideas.
Greetings and best wishes for the future!
LikeLiked by 3 people
Hi Craig. Very long discussions seeded with this comment. But at the root of the disagreement is always: physicalism. I will offer simply one observation. It is no less a positing on your part, or the parts of materialists, than what you ascribe to me, when you say I point to extra-material phenomena (or ontology, or even causation). It is absolutely an assumption, a positing, on the part of physicalism that all things, including life forms are 100% explicable, eventually, in purely material terms. So… it is a meataphysical position, not a scientific one. And I can think of no possible scientific experiment (as science, biology, physics and so forth are presently and since the 1600s constrained) which would even address this positing. It is an article of faith. And before 1600 or so, it was not, it was quite the opposite. For my asthetics, better to do science, in hopes of reaching a less biased and more complete knowledge picture of all things, without this assumption at all. Cheers to you. 🙂
LikeLiked by 2 people
Hello Rob,
I think I can appreciate something of your perspective, although for reasons of faith or otherwise, I continue to seek explanations for biological phenomena in physical terms. I hope you may be interested to read and comment on some future posts, dealing with topics such as innate human mental proclivities and their influences on human treatment of the biosphere, human emotional responses to the natural world, and issues to do with the development of environmentally sustainable economic systems
LikeLiked by 2 people
Hi Craig, yes, sure. I will watch for these future thoughts of yours. The situation between us is not personal — one can always appreciate a fellow inquisitive nature. But as one gets older it is revealed as more and more important to devote one’s time to where priorities lie. For me, I see the physicalist mindset as a key linchpin, which if exposed to a more open perspective, would have the power to release the floodgates currently obstructing human cultural potential worldwide. The materialist conception is deeply insidious; so it is extremely difficult to get most people to admit it is in fact an assumption rather than some obvious default observation. Much of my work is devoted to subtly tugging against this assumption, be it with prose or poetry.
Interesting what you mention about human proclivities vis a vis the living earth. I do not know if the proclivities you think of are innate — I would initially resist this position. But we certainly are dropping the ball regarding stewardship of our biosphere. I would immediately note that it is quite likely that materialism, as an outlook, again has its fingerprints on this particular issue. If natural resources and species are merely regarded as ‘things’, collations of atoms and sub-atomic particles and so on, then it becomes much easier to approach life from a stance of being inured to the way we treat such things. Would be interesting to see why you feel certain attitudes are innate. I would mention here that if something like evolutionary psychology is involved in the reasoning, I find the contributions and underlying concepts around EP to be extremely suspect.
Environmentally sustainable economic systems: bravo. Let such systems also include socially and humanly sustainable and I am on board and eager to hear about them. Cheers, Craig.
LikeLiked by 3 people
What a wonderful informative post and those pictures are so beautiful too!
LikeLiked by 2 people
Thank you for coming to my blog. But I don’t believe in evolution. It’s a theory. None of it makes sense. As an example, the big bang. They never explain how it came to be. You can’t get something from nothing. Creation makes much more sense. Many things have been proven from the bible. And what hasn’t been proven may have been destroyed or something else. Take care.
LikeLike
Thank you for taking the time to read our post. I hope you will also read some forthcoming posts on this site. Two of thse are to be book reviews that will have little or nothing to do with evolution, but with appreciation of nature and human use of materials throughout history.
LikeLiked by 2 people
Thank you. I see that you do indeed have other writings which is fine too,
LikeLiked by 2 people
Hello! Thank you for your comment. Please be informed that the layperson’s use and understanding of the word “theory” is very different from the same term used in modern science. For your convenience and edification, SoundEagle🦅 has included herein the Wikipedia entry of the term as follows:
In recent years, even the Catholic Church has had to accept evolution, though on a theistic basis. As for the pitfalls and fallacies of the design argument, visit the following:
http://www.iep.utm.edu/design/
http://www.pbs.org/wgbh/nova/evolution/intelligent-design-trial.html
It will be nearly or altogether impossible to claim or prove that (the theory of) evolution is wrong or invalid, for it has been estimated that if evolution (both macro and micro) were wrong, then more than 99% of all scientific disciplines would be wrong too due to the high degree of cross-collaborations and confluences of data. That is not (just) SoundEagle🦅’s claim; and it is from some scientists who have made the interconnections and stocktaking of disciplines and knowledges. When creationists try to debunk certain parts and/or the whole of the findings of evolutionists or evolutionary scientists, they have cited certain problems with some scientific claims and/or techniques which rely on or are founded on mathematics, measurements, instruments, various disciplines and so on in very interconnected ways, and have been reliably used for a long time. For example, many instruments rely on the veracity and reliability of quantum mechanics, electronics and electrical engineering, which in turn rely on other disciplines such as physics, mechanical engineering, optics and so on… It is a very highly interconnected web.
By “cross-collaborations” (whether by design or by accident, whether independently or co-dependently, and whether concurrently or not), SoundEagle🦅 meant the cumulative results, benefits and synergies from the convergence of evidence from diverse disciplines and researchers who may or may not be collaborating and/or aware of each other’s findings and activities in the first place; and SoundEagle🦅 also meant that research(ers) on/in evolution and evolutionary sciences have relied and benefited, both directly and indirectly, fertilizations, findings, paradigms and techniques from diverse disciplines. Let SoundEagle🦅 quote Michael Shermer from his essay entitled “A skeptic’s journey for truth in science” as further examples:
Yours sincerely,
ჱܓSoundEagle🦅
LikeLiked by 3 people
A very thorough mini-post that clearly explains the nature of of a scientific theory and outlines the strengths of the theory of evolution.
LikeLiked by 1 person
The fundamental law cannot be overlooked. Dogs remain dogs, cats remain cats, plants remain plants. Those do not and cannot change period. Take care now.
LikeLike
Awesome post. Thank you for sharing. Blessed are those who see plants and insects coevolve.🐝🌻
LikeLiked by 3 people
We are grateful that you took the time to read our post. You may be interested in others on this site (by SoundEagle alone) and some forthcoming joint posts.
LikeLiked by 2 people
Thank you. I will for sure take a look at the other posts. 🌺🐝
LikeLiked by 2 people
I have written a short poem about the wonderful way a flower and a bumble bee 🐝 co evolve. You may be interested to find it on my blog.
It goes like this-
How to tell you
How to tell you what is sweeter than a drop of honey
Sweeter than a grain of sugar
Sweeter than the juice of a strawberry 🍓
How to tell you
Nothing is sweeter than your tongue
Melting like ice in my mouth
How to tell you what is a flower without a bumblebee
Or a bumblebee without a flower
Or….
LikeLiked by 3 people
Thank you, Anita Bacha, for your visit, comments and compliment, as well as for sharing your lovely poem entitled “How to tell you ”. In the version of your poem shown in your comment above, SoundEagle🦅 can see that you have spiced up the poem with the two emojis 🐝 and 🍓, which are duly noted and appreciated here.
”. In the version of your poem shown in your comment above, SoundEagle🦅 can see that you have spiced up the poem with the two emojis 🐝 and 🍓, which are duly noted and appreciated here.
Happy May to you very soon and Happy Exploring other posts as you become even more familiar with the many special features and topics presented on this blog! Moreover, if your web browser can run Adobe Flash Player properly as it loads the contents of this website, then you will also be able to play games, solve puzzles and/or see more animations.

Yours sincerely,
ჱܓSoundEagle🦅
LikeLiked by 2 people
Happy May to you too.
I am glad you visited my blog and read the poem ‘How to tell you.’ The illustrating picture was clicked by me in Borehamwood in England. It was in the month of July. I am not too sure the insect is a bumblebee. I observed it and others for a long time with my iPhone ready to catch a picture of the ‘bee’ busy drinking the nectar of the flower in a bush of berries.
I just love nature and its populace.
🦋🌻🐞🌸
LikeLiked by 3 people
Thank you for sharing your delightful poem. Did you see the poem “Coexistence” by Cheryl KP in the post? Best wishes.
LikeLiked by 2 people
Sadly I scroll up and down several times I don’t see the poem Coexistence.
Possibly it’s not loading on my lappie.
LikeLiked by 2 people
The insect in the photo accompanying your poem is certainly a bumblebee, probably Bombus lapidarius or just possibly Bombus ruderarius. SoundEagle has requested me to ask whether you know what the flowers in this photo are: could they be hellebores?
By the way, you may be interested in SoundEagle’s recent long reply just above your first comment (21st March) explaining what constitutes a scientific theory and why the theory of evolution is soundly based.
Best wishes
LikeLiked by 2 people
Thank you for the the response Craig. Matter of fact, Borehamwood is a shooting ground that offers family a range of shooting facilities, like clay pigeon etc. I was there with my family to experience gun shooting because it demands focus and concentration.
On the side walk, I heard the buzzing of bees 🐝 🐝 and I followed them to click pictures.
I have some more pictures. The bees were very busy devouring what was left of flowers in wild bushes.
If you wish I can send them to you by wats app or email.
I thank SoundEagle for his profound interest in the 🐝
Hopefully we will go clay pigeon shooting again this summer.
Anita
LikeLiked by 2 people
Hi Craig
Hi SoundEagle
I forgot to tell you one important observation I made on the day I clicked the picture of the bumblebee (s).
We had to protect our ears because of the deafening noise of shooting. Yet the bumblebees and the flowers carried on with their romancing undisturbed by the sound of shooting.
They were peaceful and happy.
🌸🐝🌼🐝🌻🐝🌼🐝🌸
LikeLiked by 2 people
Hello Craig, SoundEagle, I have posted a few pictures of our bumblebee(s). Please visit my blog and see ‘Flowering’.
Happy weekend to you both.
Anita
🌸🦋🌼🌻🐝
LikeLiked by 2 people
Hello Anita,
Loud noises usually have little effect on insects, unless they drown out their auditory communications. I don’t have any personal experience of living bumblebees, as they do not occur in Australia, except for one species that was deliberately (and illegally) introduced into Tasmania in the early 1990s. I like to read accounts of exotic insects and other animals that I have not seen myself. Thank you for sharing.
LikeLiked by 2 people
Hi Craig
Thank you for the clarification.
I am truly glad and thankful to you and SoundEagle to learn so much about insects.
I have posted a few pictures on my blog. Please have a look at my latest post ‘Flowering.’
En passant, I live in Mauritius and here, there’s no bumblebee.
We have the honey bee. It’s completely black in color, whereas the bumblebee is black and orange. It also uses its long arms to embrace the flower.
🌸🐝
LikeLiked by 2 people
Pretty long post about this very interesting topic…
LikeLiked by 4 people
This is just one example of a widespread and important phenomenon. I hope you will also read our forthcoming posts on other topics.
LikeLiked by 1 person
I find it very lengthy too but very absorbing.
LikeLiked by 4 people
I too have always found evolutionary processes very absorbing. I wish I could have specialized in this field during my research career! Many thanks for your interest.
LikeLiked by 2 people
Thank you too
LikeLiked by 2 people
I like this kind of subjects, but I don’t like to long texts. I don’t have any more patience for such a long reading…
LikeLiked by 3 people
I respect your opinion.
LikeLiked by 3 people
😊 10ks!
LikeLiked by 3 people
Great post!
LikeLiked by 2 people
Thank you for your interest. SoundEagle is now trying to restore the images that have been lost from the Gallery section. Best regards.
LikeLiked by 3 people
Great!
Thank you very much
LikeLiked by 2 people
Thank you Craig, I now see the poem Co-Existence of Cheryl KP 2017.
Awesome 🌹🐝❣️
LikeLiked by 2 people
As I mentioned before the fundamental law states that plants remain plants, insects remain insects and so on. Noting evolves at all that’s why you can’t prove otherwise no matter what so-called science you base your claims on. Take care now.
LikeLike
I am sorry that we cannot agree about evolution. However, I hope that you have found something of interest in our post, and wish you all the best for the future.
LikeLiked by 1 person
I can assure you my dear that my friend, walking companion, and confidant that has never once reveled a secret is not a dog.
Without a doubt Michelangelo created the masterpiece called the “Mona Lisa”, but he never mucked with it afterwards. If it is perfect at conception what tweaks can make it better?
LikeLiked by 2 people
I love and sometimes fear the true interconnectedness of it all and how I am stuck to the outcome of what others might be doing, but also thrilled at the glue that binds me to the butterflies and their habitat we saved from being chopped down around the local train station.
LikeLiked by 3 people
My apologies for the very late reply. The interconnectedness of life is an important lesson for all of us, and something that we disregard to our great peril. I am pleased that we share an interest in conserving the fragments of the living natural world that still remain. Thank you for taking the time to read this post; I hope that you will find future posts on this website to be of equal interest.
Best regards
LikeLiked by 2 people
The write-up is obviously very interesting, educative since it elucidates the inter-connection between different species and above all well researched. There is a connect between all the creations of the Mother Nature – one is incomplete without other. You have really given everything which essentially keeps the interest of the readers ticking. A must read write-up for information on correlation between all which is created by our CREATOR! Just wonderful to say the least. Regards.
LikeLiked by 2 people
Hello Harbans! Thank you for your comment and compliment, especially with respect to your recognizing the interconnections of all lives on Earth.
SoundEagle🦅 has already notified Dr Craig Eisemann, the co-author of this post, of your freshly composed comment so that he may give you a reply at his earliest convenience.
Since you have mentioned twice that you are going to read and comment on the special post entitled “Khai & Khim: For Always and Beyond Goodbye”, SoundEagle🦅 is somewhat caught by surprise that you read and comment on this post first. You must have been very curious about or keenly interested in natural science in general as well as evolutionary biology and ecology in particular.
Looking forward to interacting with you in the said post published at https://soundeagle.wordpress.com/2019/08/31/khai-khim-for-always-and-beyond-goodbye/
 Yours sincerely,
Yours sincerely,
ჱܓSoundEagle🦅
LikeLiked by 1 person
Hello Harbans; I am pleased that you found our efforts to elucidate this topic interesting and useful. The intimate relationships between, and the coadaptations of, insects and plants have long fascinated me and others, and I am sure that much remains to be discovered about the interactions between these vast and diverse groups of organisms. It is a tragedy that many of these species may be lost to the earth through human-caused extinction before they are appreciated by or even known to us.
Please accept my best wishes and invitation to read future posts on this website.
LikeLiked by 2 people
My gratitude to you for your comments. In fact, we humans consider boundaries within which we live and interact an important one and leave all other creations by our Creator thereby losing the proximity of our approach to birds, plants, insects etc. and their inter-connection for enhancing and growth of echo-system which directly and indirectly help the humans. We are on the burning spree the jungles as has been happening in Amazon forests. How many of the species of plants, animals and birds have been lost only the CREATOR knows the best. These are not merely the ones which live overland and what about the ones which live by making burrows or holes for their living places by taking out earth or those birds living in their nests. Or other razing fires or the creatures living in the oceans; which are also dying due to throwing poisonous left-overs after our use in the seas.
I was brought up in a village which was near the forest area and there we used to go and enjoy every bit of songs which the Mother Nature created for us. I think, that was a good life when I used to run after a cat or dog or a rat, or a parrot, or played with plant like touch-me-not? We have now lost contact with all these and that is why we are suffering the pangs of different diseases which were not there when people used to befriend nature rather than cutting forests for habitation or burning forests for agriculture etc.
What a life it would be
if we care not merely for ourselves
but everything that our Lord has made
but our greed and ego
just spurs us to enhance our domain
at the cost of our fellow creations
Loving animals, birds, flora fauna
is always reciprocated
lets make friend with anything and everything that Nature contains in abundance
Cherish and enjoy it
All these will give us eternal joy
which we cannot get from any materials
which we enjoy for a moment
and then forget.
With regards,
HARBANS
LikeLiked by 2 people
I much appreciate your heart-felt observations on the destructive results of the rampant and seemingly unstoppable “human enterprise” that seeks to convert ever more of the natural biosphere to human material use. Being in what nowadays is the somewhat unusual situation of again living in the house in which I grew up, I am in a position to see clearly the changes in the climate and in the composition and abundance of the surrounding fauna that have occurred as a result of human activities over a period of more than fifty years. You may also be interested in some forthcoming posts on this site, which attempt to address what is happening to the natural world and to explore some of the innate characteristics of human nature that influence the ways in which humans behave towards the living biosphere.
Once again, thank you for your interest and support.
LikeLiked by 2 people
My heartfelt thanks for your inspiring commentary.
Factually, some of us are spoiling the echo-system, thereby spoiling the balance of nature, by so doing, the common man is suffering.
The contribution of good writers and well-wishers work for the well-being of the Mother Nature thus proving counter-productive. What is happening now in our cities and towns due to pollution impacting our health.
I shall go through your future write-ups for learning and adding to my knowledge.
WITH WARM REGARDS
HARBANS
LikeLiked by 2 people
I hope that our forthcoming posts will be informative and interesting for you, and I look forward to interacting further with you when you have had an opportunity to read them. Please accept my best wishes for the future.
LikeLiked by 2 people
Craig sir thanks a lot for informing me about your next post. I shall visit your blog and add to my know-how about our Creator’s creations which you assiduously research and bring to our domain.
I heartily reciprocate the wishes. God may bestow you strength and fortitude for educating us still more.
REGARDS
LikeLiked by 2 people
Do you hav bees
LikeLike
I believe so, there is an inherent symbiosis in nature. Balance & harmony is the result, mankind has forgotten we are a part of the whole, do not own anything, but merely stewards of this planet.
Th mystic Osho used to say, “If there were no trees then birds would not exist, conversely if the birds start disappearing then trees will too”!
We’re all interconnected. ❤
LikeLiked by 3 people
Thank you for your observations on the relevance of this question to the wider issues you raise.
The subject of this post is increasingly topical in view of the “insect apocalypse” that is becoming evident in various parts of the world, and the implications that this holds for other forms of life. Unfortunately, the majority of our species seem not to apply their characteristic capacity for empathy and rational thought consistently to the predicament in which we and our fellow inhabitants of this planet are increasingly mired, but often to indulge atavistic, irrational drives and prejudices.
We hope to consider some aspects of this issue in a forthcoming post.
LikeLiked by 3 people
two species who co-evolve are like two planets turning around each other, one may seem to turn around the other but both turn around a commun gravity point that may be or may be not inside of one of the planets
LikeLiked by 2 people
Happy mid-April to you, Gérard!
LikeLiked by 2 people
This is an interesting and thought-provoking analogy. For two interacting species, it is quite possible that Species A may have much less effect on the evolution of Species B than the reverse, if Species A is much less significant for the biology of Species B than vice versa. However, it is also likely, as your analogy suggests, that any two significantly interacting species will each have at least a slight effect, through selection pressure, on the other’s evolution, even if this is not readily apparent upon observation of the less-affected species.
LikeLiked by 2 people
I point to the vast sums being spent to obscure the effects humanity is having on all our fellow co-evolutionists.
LikeLiked by 2 people
What an interesting and very fascinating post. It’s clear you have such an in-depth knowledge and passion for this subject!
LikeLiked by 3 people
Hello Rachel! Thank you for your comment and compliment. SoundEagle🦅 will soon notify Dr Craig Eisemann, a lover of moths and the co-author of this post, of your freshly composed comment, so that he too may enjoy your lovely poem entitled “Like a moth to your flame…” as follows:
ჱܓSoundEagle🦅
LikeLiked by 2 people
Thank you, I’m so happy you enjoyed that poem. What a compliment you would think someone else might enjoy it too.
LikeLiked by 3 people
The pleasure is mine, Rachel, as the previous bespoke reply is a stylish tribute to your poetic flair and your love of moths.
Dr Craig Eisemann will duly appreciate and be impressed by both your poem and the eye-catching golden moth. Hopefully, he will respond to your comment(s) within a day or two, as he is currently occupied by a few errands.
In addition, if you were to be so kind as to leave your feedback or critique at the multimedia post entitled 🎼🎹—THE—🎹—LAST—🎹—RAG—🎹🎵🎶 featuring animated moths/butterflies, SoundEagle🦅 will once again be so obliged and delighted that another bespoke comment will be delivered to you in style.
In any case, may the music and poem there bring you some creative “distractions” or “diversions” amidst the disruptions and woes engendered by the viral pandemic! Please enjoy to your heart’s content.
 Yours sincerely,
Yours sincerely,
ჱܓSoundEagle🦅
LikeLiked by 1 person
Insect-plant coevolution is a long-standing interest of mine, although never directly a subject of my research. I’m glad that you enjoyed the results of our effort!
LikeLiked by 2 people
I liked your beautiful and profound poem, with its lepidopteran imagery, and also appreciated the spectacular photograph of what I believe is a Polyphemus moth, native to much of North America, that accompanied this poem on youir website. Since early childhood, I have been fascinated by the subtly beautiful colouration of many night-flying moths, and this species provides a wonderful example. I was wondering whether you could tell me the source of this photograph.
LikeLiked by 2 people
I will gave a look, it was a beautiful photograph. Thank you for your very kind words.
LikeLiked by 2 people
Really detailed and sound research!
LikeLiked by 2 people
Coevolution is a common mechanism in nature because it presents many advantages to the organisms (but also comes with strings attached). It’s a way to avoid competition (https://bit.ly/316qlDH), find a niche and coexisit. Your example of V. cornigera avoiding competition because of their relationship with ants perfectly illustrates this. The science is fascinating and very extensive. I am so happy that I have delved so deep into the topic (I don’t know if you are an ecologist or environmental scientist. If you aren’t this is even more impressive!)
Great job again, SoundEagle 😀
LikeLiked by 2 people
Another bespoke acknowledgement is due to you for perusing and commenting on this expansive post, in which you have shown a considerable interest. Thank you for your complimenting SoundEagle🦅 (and by extension, Dr Craig Eisemann), on the quality and magnitude of the research effort entailed in producing the work, which in parts has even ventured far beyond the purview of answering the research question to include a detailed summary of the academic environment and corporate culture facing contemporary scholars.
In resonating with Dr Craig Eisemann who has promptly replied to your comment, SoundEagle🦅 would like to inform you belatedly that this post has a tripartite structure comprising 🥀🦋 ❀ Entrée ❀ 🐝🌺, 🌿🐜 ✿ Main Course ✿ 🐛🍃 and 🌻🐝 ❀ Desserts ❀ 🐞🍂. SoundEagle🦅 is responsible for the Entrée and Desserts, whereas Dr Craig Eisemann for the Main Course.
Given your professional background and interest, you are very welcome to make further perusals and comments here as you inspect and ingest the three-course offering to your heart’s content.
Speaking of insects, the musical and multimedia post entitled 🎼🎹—THE—🎹—LAST—🎹—RAG—🎹🎵🎶 features animated moths/butterflies plus music and poem to bring you some creative “distractions” or “diversions” amidst the disruptions and woes engendered by the viral pandemic. Please enjoy.
Yours sincerely,
ჱܓSoundEagle🦅
LikeLiked by 1 person
Saurab Babu, I am glad that you appreciated our efforts! I have studied ecology at undergraduate level (many years ago), but chose insect behaviour and sensory physiology as my field of research, and later moved on to participate in an unsuccessful attempt to develop a vaccine to protect sheep against myiasis-producing blowflies. SoundEagle (who wrote the long introduction to this post) does not have a comparable background in biology, but having already mastered several diverse fields of knowledge, was well able to research and compose this contribution.
I have visited your website, and am impressed both by your aspiration to provide education in a range of environmental sciences, and by your achievement in creating a diverse and well-presented learning resource. I wish you every success for your vital endeavours in the future.
LikeLiked by 1 person
You have managed to collect a TREMENDOUS amount of information on coevolution, SoundEagle! I find the relationship between plants and animals fascinating. For me, the beauty of the natural world reflects God’s own. Thank you for the education. Stay safe and well. 🙂
LikeLiked by 2 people
Hello Anna,
We are grateful that you have read and appreciated our efforts. I began my research career with a study of insect responses to stimuli (mainly chemical) from their host plants and have been interested in insect-plant relationships ever since, although my research subsequently took other directions.
My contribution to this post was confined mainly to writing the text and finding some illustrations for the second part, while SoundEagle wrote the long introduction, found many additional illustrations (some not presently accessible, unfortunately) and was responsible for the design and layout of the whole post.
We wish you all the best for the future, and hope that you will be similarly interested in future posts on this website.
LikeLiked by 2 people
🥀🌺Plants 🔄 Insects🐝🦋
💖 Dear Anna 💝
SoundEagle🦅 is delighted by your frequenting and commenting on this co-authored work, a substantial research undertaking to highlight the far-reaching ramifications of mutualism and evolutionary partnership, as much as this post has been the result of a similar kind between two intellectuals bound by enduring friendship and comparable interests.
This post has a tripartite structure comprising 🥀🦋 ❀ Entrée ❀ 🐝🌺, 🌿🐜 ✿ Main Course ✿ 🐛🍃 and 🌻🐝 ❀ Desserts ❀ 🐞🍂. SoundEagle🦅 is responsible for the Entrée and Desserts, whereas Dr Craig Eisemann for the Main Course.
Please feel free to make further perusals and comments here to your heart’s content as you savour the three-course offering.
May you feel Younger than Springtime in September!
ჱܓSoundEagle🦅
LikeLiked by 3 people
Wow I love this 😍
LikeLiked by 2 people
We are pleased when we can provide a pleasurable, as well as an educational, experience for our readers. Both authors increased their knowledge considerably in preparing their respective contributions to this post. It was a new departure for SoundEagle, who does not have a formal background in biological sciences, and while I have worked professionally as a biologist, evolutionary biology, although a long-standing interest of mine, was never part of my research.
We apologize for the fact that many of the images originally included in the Gallery section are currently not available.
Thank you for your interest in, and attention to, our work.
LikeLiked by 1 person
Thank you and keep up the great work!
LikeLiked by 2 people
Hey, Sound Eagle. Have a blessed rest of the summer!
LikeLiked by 2 people
Useful content and well written by a maestro
LikeLiked by 2 people
We are glad that our work has been of use to you, and are grateful for the compliment.
LikeLiked by 1 person
Another endlessly intriguing post. As I was once upon a time a biology major, the topic of co-evolution holds great interest for me. You, SoundEagle, and Craig Eisemann are excellent collaborators!
LikeLiked by 3 people
Fascinating. Amber ❤🙏
LikeLiked by 2 people
As co-author of this post, I am pleased that you found it to be of interest, and hope that you will also enjoy some forthcoming posts on this website.
LikeLiked by 2 people
This is a very informative post. Thank you for liking so many of my comments.
LikeLiked by 2 people
It seems as if the answer to the question “Do Plants and Insects Coevolve?” is far from a simple “yes” or “no.” There are many factors to consider and many distinctions to be made in understanding the question and its answer. I appreciate your care and diligence in exploring them.
Although it may not be apparent from the title, this post also offers a wealth of information for orchid lovers.
I just wanted to let you know that many of the images are currently replaced by “Image not found”:
“Orchid (Dracula lafleurii) flower with pollinator flies, Ecuador”
“King Spider Orchid (Caladenia pectinata) being visited by male parasitic wasp pollinator which is attracted to the flower by faux female wasp pheromone, Australia” (and many subsequent)
“Pair of marsh fritillary butterflies (Euphydrayas aurinia) resting on a heath orchid flower (Dactylorhiza fuchsii) in the countryside of Devonshire” (and subsequent)
LikeLiked by 2 people
On the balance of accumulated data and empirical research in evolutionary biology, the answer seems to lean quite substantially towards the affirmative. Thank you for complimenting SoundEagle🦅 (and by extension, Dr Craig Eisemann) on the care and diligence in exploring the “many factors to consider and many distinctions to be made in understanding the question and its answer”.
Perhaps your own interest in the question “Do Plants and Insects Coevolve? 🥀🐝🌺🦋” is in no small part due to the recent publication of your trilogy entitled “Plant Intelligence and the Imaginal Realm” comprising “Part I—Perceiving Gaia”, “Part II—Making America Great Again” and “Part III—Learning to Trust Yourself”.
Please be informed that this post has a tripartite structure comprising 🥀🦋 ❀ Entrée ❀ 🐝🌺, 🌿🐜 ✿ Main Course ✿ 🐛🍃 and 🌻🐝 ❀ Desserts ❀ 🐞🍂. SoundEagle🦅 is responsible for the Entrée and Desserts, whereas Dr Craig Eisemann for the Main Course.
SoundEagle🦅 has been aware of the missing images for more than a year or two, and will endeavour to resurrect them once a certain post composed by Dr Eisemann is fully formatted and published on this website by SoundEagle🦅 later in the year or early next year.
Given your professional background and interest, you are very welcome to make further perusals and comments here as you inspect and ingest the three-course offering to your heart’s content. Furthermore, SoundEagle🦅 is really curious to know what you thought of the digression into a detailed summary of the academic environment and corporate culture facing contemporary scholars.
Speaking of insects, the musical and multimedia post entitled 🎼🎹—THE—🎹—LAST—🎹—RAG—🎹🎵🎶 features animated moths/butterflies plus music and poem to bring you some creative “distractions” or “diversions” amidst the disruptions and woes engendered by the viral pandemic. Please enjoy them in full.
Your observation or conclusion that “this post also offers a wealth of information for orchid lovers” is appreciated. Moreover, you will be pleased to know that you can assess four other plant-oriented websites created and managed by SoundEagle🦅:
ჱܓSoundEagle🦅
LikeLiked by 1 person
Reblogged this on worldtraveller70.
LikeLiked by 2 people
Reblogged this on worldtraveller70.
LikeLiked by 2 people
[…] Source: Do Plants and Insects Coevolve? 🥀🐝🌺🦋 | SoundEagle 🦅ೋღஜஇ […]
LikeLiked by 2 people
A wonderful read, and a thoughtful exposition, much for me to ponder.
LikeLiked by 2 people
We are glad that you appreciated our efforts, and as always are pleased that this post stimulated your thinking about the information presented and its implications.
LikeLiked by 3 people
It sure did.
LikeLiked by 2 people
species have hollow thorns and pores at the bases of their leaves that secrete nectar (see image at right). These hollow thorns are the exclusive nest-site of some species of ant that drink the nectar. But the ants are not just taking advantage of the plant they also defend their acacia plant against herbivores.
LikeLiked by 1 person
What an interesting question. Thanks for making me think!
LikeLiked by 2 people
Thank you once again for your amazing productivity, creativity and research. Um… what do you do in your spare time? 🙂
I am horrified by those Orchid hunters. What greed can do!
Since humans and insects have coevolved, and plants and insects have coevolved, and humans and plants, it seems to me that one could write a pretty good nursery rhyme with this theme.
🙂
Bob
LikeLiked by 2 people
Hello Bob,
As SoundEagle’s co-author on this post, I am grateful for your attention to our efforts.
It would be interesting if you could devise a nursery rhyme on your suggested theme and contribute it here.
I apologize for the unavailability of most of the images that were originally in the Gallery; we have not yet found the time to replace them.
Best wishes
LikeLiked by 2 people
This is an extensive and well researched post on co-evolution. There are some really nice examples highlighted in terms of pollination and seed dispersal mutualisms. And a great gallery of resources to draw on for further reading. Certainly leaves open other options for topic areas in the biological sciences – is runaway sexual evolution next?
LikeLiked by 2 people
Thank you for taking the time to peruse this post and to compliment SoundEagle🦅 and Dr Craig Eisemann here. Also appreciated is your acknowledgement on LinkedIn that “[t]he nice thing about [this] site is the in-depth treatment of topics”, based on your observation or conclusion on the quality and features that you have so far experienced at the website.
Circumventing the many restrictions and conventions imposed by academic journals whose contents are often hidden behind paywalls, self-publishing certainly presents numerous advantages, which, when combined with the dynamic nature of a blog post offering contents that can be freely viewed and shared by visitors and commenters, are attractive to the likes of us who have been inveterate teachers and communicators. Accordingly, this post has a tripartite structure comprising 🥀🦋 ❀ Entrée ❀ 🐝🌺, 🌿🐜 ✿ Main Course ✿ 🐛🍃 and 🌻🐝 ❀ Desserts ❀ 🐞🍂. SoundEagle🦅 is responsible for the Entrée and Desserts, whereas Dr Craig Eisemann for the Main Course.
SoundEagle🦅 and Dr Craig Eisemann have been cognizant of the missing images for more than a year or two, and will endeavour to resurrect them once a certain post composed by Dr Eisemann is fully formatted, illustrated, animated and published on this website by SoundEagle🦅 later in the year.
Given your professional background and interest, you are very welcome to make further perusals and comments here as you inspect and ingest the three-course offering to your heart’s content. Furthermore, SoundEagle🦅 is really curious to know what you thought of the digression into a detailed summary of the academic environment and corporate culture facing contemporary scholars.
This post belongs to the category entitled Facing the Noise & Music, under which the following posts have also been classified and listed in reverse chronological order:
Happy mid-September to you!

ჱܓSoundEagle🦅
LikeLiked by 2 people
Thanks Sound Eagle
Yes I agree blog posts are good way to get academic ideas out there, but particularly to summarise thoughts for a more general audience. Which can also be an important synthesis of ideas and thoughts, which may be weeded out in the submission of a full academic paper.
Publishing in the academic literature does however make sure that article are independently referred and so they remain the quality milestone of new work.
Blog posting and academic literature publishing can and should go hand in hand though.
LikeLiked by 2 people
We are more or less in agreement here. Apart from the accessibility of academic works as mentioned earlier, there are many other extant issues that remain to be addressed. Academic papers published in most if not all conventional journals tend to be highly limited in scope, format and feature, mostly presented in monochrome or greyscale and rarely in colour; cannot include large or complicated illustrations, maps, charts, schematics and/or statistics; unable to incorporate audiovisual contents, videos, animations, pop-ups, hyperlinks, comments, revisions, extensions, user interfaces and other real-time, dynamic or interactive components, let alone social media integration; and ill-suited or ill-prepared to deal with highly innovative, interdisciplinary and/or multidisciplinary works, not to mention that there continue to be few or no referees and reviewers qualified or knowledgeable enough to evaluate them properly and cogently without intellectual oversight and bias.
Let us examine just one example regarding pop-ups. In this website, tooltips, a form of pop-ups, also deliver full bibliographical information. Considering that certain 📑Posts and Pages📃 are very long and extensive, the provision of tooltips as a way of implementing citations using the combination of on-demand, in-situ pop-up references and hyperlinks is a matter of prudence and practicality to dispense with the overly expansive space that would otherwise be required to accommodate a bibliography or reference list whose entries are in the hundreds, which would have made any one of the already lengthy 📑Posts and Pages📃 even longer.
Other insidious matters regarding conventional publication of academic papers have also been discussed by SoundEagle🦅 in a number of academically written posts. The following are just a few chosen excerpts to highlight some of these matters. The first set is taken from the multidisciplinary book-length post entitled 💬 Misquotation Pandemic and Disinformation Polemic: 🧠 Mind Pollution by Viral Falsity 🦠 published with multipronged analyses under the following categories: Behavioural Science, Cognitive Science, Critical Thinking, Cultural Studies, Environmentalism, Epistemology, Ethics, Evolutionary Biology, Evolutionary Psychology, Facing the Noise & Music, History, Human Nature, Information Science, Journalism, Logic, Media Studies, Philosophy, Political Science, Psychology, Quotes, Science, Social Media and Social Science:
The second set is sourced from an equally multidisciplinary but even lengthier post entitled The Quotation Fallacy “💬”:
There is a decent, illuminating account from a prolific academic by the name of Peter Cameron, who, like you, also maintains a blog. After having decades of experience in academic publication, he has decided to write a detailed summary and stocktaking in a post entitled “Publication: an author’s view”, in which he concludes as follows:
Looking forward to be enlightened by your thoughts and experiences on these matters!

ჱܓSoundEagle🦅
LikeLiked by 2 people
Hello Prof. Lowe,
Apologies for the late reply. The next post in which I have some involvement will bear some relevance to (human) evolutionary biology and psychology, but regrettably none to sexual evolution, human or otherwise.
Evolutionary biology in general is a long-standing interest of mine, and insect-plant relationships, particularly insect sensory physiology and behaviour as it relates to selection of host plants, constituted my original research speciality before I turned, of necessity, to other fields of research.
Both co-authors of the present post learned a good deal about insect-plant coevolution while preparing for it, and it is just possible, although not likely, that we may return to this topic at some time in the future.
Please accept my best wishes and thanks for the interest that you have shown in our work.
LikeLiked by 1 person
How fascinating! I never really thought about coevolution before, but of course it makes perfect sense. Thanks for teaching me about this interesting topic. : )
LikeLiked by 2 people
Iím amazed, I have to admit. Seldom do I encounter a blog thatís equally educative and interesting, and let me tell you, you’ve hit the nail on the head. The problem is something that not enough people are speaking intelligently about. Now i’m very happy that I came across this in my hunt for something regarding this.
LikeLiked by 1 person
An interesting discussion is worth comment. I think that you ought to publish more about this subject, it may not be a taboo subject but usually people do not discuss such topics. To the next! Many thanks!!
LikeLiked by 1 person
You’re so awesome! I don’t think I have read through anything like that before. So great to discover someone with some unique thoughts on this issue. Really.. thank you for starting this up. This site is one thing that’s needed on the internet, someone with a bit of originality!
LikeLiked by 1 person
[…] 2016 […]
LikeLiked by 1 person
[…] Continue to find out more: Do Plants and Insects Coevolve? […]
LikeLiked by 2 people
Like lots of other articles of yours very interesting, though this time there were many pictures not available to be seen. Perhaps also something to think about (for the future) is to split the articles in several postings, so that you can catch more people who will be able to read the posts in parts at times when they have time, so that they would not click it away after some minutes of reading, without having it fully read..
LikeLiked by 2 people
Thank you for your interest in our post. The authors are aware that many images, especially in the Gallery, have not been available for some time, and hope to address this in the near future.
As regards splitting of articles, both authors are inclined to think that, given the fairly detailed menus that are provided at the beginning and end of long posts, readers could conveniently resume reading from where they had left off on an earlier occasion. This arrangement allows greater cohesion and connectivity among parts of posts that are intended to be grasped as a whole, rather than as a series of disconnected aspects of the topics that are dealt with.
Please accept my apologies for the late reply.
LikeLiked by 1 person
Thank you very much for your reply. Concerning the lateness of the reply, do not worry, better a reply than none. We, too, have to face hundreds of e-mails per day, and follow several blogs and articles, so that it often can take some time before they got answered or that we react on some writing.
LikeLiked by 2 people
Excellent post. This is something I had not really thought about before.
LikeLiked by 2 people
Thanks for the post. It was highly informative and a pleasure to read. Have a nice day!
LikeLiked by 2 people
[…] askedDo Plants and Insects Coevolve? 🥀🐝🌺🦋it is time to […]
LikeLike
Hi SoundEagle, one more aspect: I think that all organisms which have sexual reproduction coevolve with themselves, or rather, different genes that can be recombined in sexual reproduction (or even parts of genes that can be recombined in crossing over events) co-evolve. We could think of those genes or parts of genes as species that together produce a phenotype and co-evolve. Such a co-evolution might start with a week coupling between them and then they could together “zoom in” on a novel solution to an evolutionary problem.
I had this idea when I read an article (many years ago, I think in Scientific American, but I don’t remember exactly) about wind polinated plants, where the shape of the femal flowers and the shape of the pollen caught by them literally filtered out each other in a process of co-evolution of different parts of the same genome. So features within one species that fit together can amplify each other and lead to a new combined phenotype, leading to features of the phenotype in which several or many genes contribute. I even think that learned features might become part of such a phenotype (e.g. a learnt route of a migratory bird might become a factore that filters out genes contributing to its “hard-wiring” in the brain, as a kind of “virutal phenotype” without a genetic basis). This “learned phenotype” could then become part of the phenotypical coupling of a group of genes.
(I noticed you liked some of my comments on different web sites. I am planning to restart blogging “soon” (sometime next year, probably)(, on wordpress or elsewhere, and I will let you know then since you seem to be interested. Meanwhile, you can look for some articles I wrote on the journal “Borderless Philosophy”. Look there fore Andreas Keller (in the editions BP2 (2019) to BP5 (2022), I have contributions in all four editions.)
LikeLiked by 2 people
Dear Nannus,
This is an interesting and plausible idea. Certainly, genes within a population could be selected according to how effectively they complement each other to enhance the fitness of the phenotype. Such a process could lead to a gradual refinement of parts of the organism in relation to their function as organs (for example) and as components of a viable integrated whole, as natural selection drives genetic and phenotypic change towards maximal efficiency and hence fitness of organisms within the population.
I note your suggestion that learned behaviours (such as following specific migration routes) could initiate a genetic selection process whereby they eventually become encoded in the genome. This idea seems consistent with the theory of natural selection, and so is also a valuable and plausible contribution.
Best wishes
Craig Eisemann
LikeLiked by 2 people
[…] – The co-evolution of plants and insects https://soundeagle.wordpress.com/2016/08/17/do-plants-and-insects-coevolve/ […]
LikeLiked by 2 people
[…] Do Plants and Insects Coevolve? 🥀🐝🌺🦋 […]
LikeLike
Ha You used up all my reading time for the day! Knowing the lifecycles of insects got me my first post college job poisoner. What are your thoughts on the effects of micro poisons such as Pyrethrins and CO2. got to run
LikeLiked by 2 people
Thank you for taking the time to read our post.
Pyrethrins are among the least toxic insecticides to mammals, and natural pyrethrins, at least, quickly break down in the environment. However, I think that the use of all insecticides should be kept to a minimum. Most of my research career was devoted to studying possible alternatives to insecticides for pest control.
I’m not sure that I understand your meaning in including carbon dioxide in this context. Would you like to elaborate on this point?
LikeLiked by 2 people
I have to assume your research into alternatives was motivated due to the sub-acute effects of toxins in the environment. CO2 is a sub-acute toxin that has doubled in the environment in the last 50 years. In higher organism this rapidity hard to absorb and I wonder if the effects on brain chemistry might be similar to Pb and Pyrethrins. I speculate that this might explain the uptick in violence.
LikeLiked by 2 people
My research into alternatives to insecticides was motivated by both ecological concerns (effects on non-target organisms) and the possibility of as yet unknown sub-acute effects on humans.
I do not think that, in most situations, ambient levels of carbon dioxide (at present, an average of just less than 420 parts per million in atmospheric air) are a cause for concern, at least as regards direct effects on human health. For comparison, exhaled human breath contains about 38,000 parts per million of carbon dioxide as a result of normal metabolic processes. Effects of this gas on global climates and the acidity of the oceans are a different matter, of course.
LikeLiked by 1 person
Darwin’s insights have always captivated me, especially his identification of carnivorous plants as a pioneering discovery. From a young age, I’ve cultivated numerous types of these fascinating plants. My lifelong passion for botany has led me to study the field extensively. Currently, my home’s landscape is adorned with hundreds of hydrangeas. Growing up in a family that farmed around 50,000 acres, the principles of cohabitation and coexistence were fundamental to our approach. Within the realm of soybeans, the process of nitrogen fixation is facilitated by the bacterium Bradyrhizobia japonicum. This symbiotic relationship allows the plant to acquire nitrogen, while the bacteria gain the energy necessary for their growth and development.
LikeLiked by 2 people
Thank you for submitting your very first comment to this academically written post, which has a tripartite structure comprising 🥀🦋 ❀ Entrée ❀ 🐝🌺, 🌿🐜 ✿ Main Course ✿ 🐛🍃 and 🌻🐝 ❀ Desserts ❀ 🐞🍂. SoundEagle🦅 is responsible for the Entrée and Desserts, whereas Dr Craig Eisemann for the Main Course. Given your background and interest, you are very welcome to make further perusals and comments here as you savour the three-course offering to your heart’s content. Furthermore, SoundEagle🦅 is considerably curious to know what you thought of the extensive digression into a detailed summary of the academic environment and corporate culture facing contemporary scholars in relation to your own experiences in these regards.
Speaking of insects, the musical and multimedia post entitled 🎼🎹—THE—🎹—LAST—🎹—RAG—🎹🎵🎶 features animated moths/butterflies plus SoundEagle🦅’s original music and poem to bring you some creative “distractions” or “diversions”, given that you like listening to music and appreciating poetry. Printable music scores and high-resolution videos are also available to you there.
Your observation or conclusion that “Darwin’s insights have always captivated [you], especially his identification of carnivorous plants as a pioneering discovery” as well as your “lifelong passion for botany” are both appreciated and admirable. Given your penchant and orientation, you will be pleased to be informed that you can assess four other plant-oriented websites created and managed by SoundEagle🦅:
There has been no shortage of scientists who are or were religious. Moreover, regardless of the degree to which some scientists are religious or atheistic, it is very unfortunate that too often even those who claim to believe in and adopt the scientific method still cherrypick the data and refuse to examine contrary evidences. They fail to understand and address many valid points, perspectives, domains and dimensions, and hence it is impossible for them to evaluate and change their standpoints, approaches and behaviours. You might have heard of this quote:
Indeed, there are evolutionary bases in people’s sense of morality and in their behaviours as well as in their religiosity. You will find a great deal of new understandings in multidisciplinary fields such as sociobiology, evolutionary psychology and behavioural sciences, epigenetics, brain and cognitive sciences, gene-culture coevolution, and many more. The following is an extensive and analytical post with a catchy title. It provides a multidisciplinary analysis and overview regarding human nature and other salient matters harbouring profound implications and existential ramifications. SoundEagle🦅 is hereby recommending it to you insofar as it deserves to be perused and critiqued by someone of your intellectual calibre and philosophical persuasion. For your convenience, clicking the large title below will instantly transport you to the said post:
ჱܓSoundEagle🦅
LikeLiked by 1 person
Dear Kenn,
As an author, it is always gratifying for me to attract the interest of people who have an enthusiasm for the natural world and biological processes. The example that you gave of a symbiotic association between the soybean plant and Bradyrhizobium is an excellent illustration of a coevolutionary relationship, in this case between a plant and a bacterium.
I have a long-standing interest in the relationship between plants and phytophagous insects, and my earliest research was concerned with an aspect of this.
Best wishes for your present and future botanical studies.
LikeLiked by 1 person